
Long-Term Persistence of Mitochondrial DNA Instability in HIV-Exposed Uninfected Children during and after Exposure to Antiretroviral Drugs and HIV

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Study Population
2.3. Sample Collection and Processing
2.4. Detection and Quantification of Mitochondrial DNA Instability: The eKlispe-v2 High-Throughput Computational Pipeline
2.5. Statistical Analysis
3. Results
3.1. Study Population Characteristics at D7, W50, and Y6
3.2. MtDNA Alterations among HEU
3.3. Determinants of mtDNA Instability at D7
3.4. Impact of the Prophylactic Drug Exposure on mtDNA Instability
3.5. Comparison of Clinical and Biological Characteristics at Y6 by mtDNA Heteroplasmy Rate Early in Life
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McBride, H.M.; Neuspiel, M.; Wasiak, S. Mitochondria: More Than Just a Powerhouse. Curr. Biol. 2006, 16, R551–R560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, J.; Baptiste, B.A.; Kim, E.; Hussain, M.; Croteau, D.L.; Bohr, V.A. DNA Damage and Mitochondria in Cancer and Aging. Carcinogenesis 2020, 41, 1625–1634. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.A.; Johnson, K.A. Exonuclease Proofreading by Human Mitochondrial DNA Polymerase. J. Biol. Chem. 2001, 276, 38097–38107. [Google Scholar] [CrossRef] [PubMed]
- Longley, M.J.; Nguyen, D.; Kunkel, T.A.; Copeland, W.C. The Fidelity of Human DNA Polymerase Gamma with and without Exonucleolytic Proofreading and the P55 Accessory Subunit. J. Biol. Chem. 2001, 276, 38555–38562. [Google Scholar] [CrossRef] [Green Version]
- Gorman, G.S.; Chinnery, P.F.; DiMauro, S.; Hirano, M.; Koga, Y.; McFarland, R.; Suomalainen, A.; Thorburn, D.R.; Zeviani, M.; Turnbull, D.M. Mitochondrial Diseases. Nat. Rev. Dis. Primers 2016, 2, 1–22. [Google Scholar] [CrossRef]
- Goldstein, A.; Falk, M.J. Mitochondrial DNA Deletion Syndromes; University of Washington, Seattle: Seattle, DC, USA, 2019. [Google Scholar]
- Kauppila, T.E.S.; Kauppila, J.H.K.; Larsson, N.-G. Mammalian Mitochondria and Aging: An Update. Cell Metab. 2017, 25, 57–71. [Google Scholar] [CrossRef] [Green Version]
- Picard, M.; Vincent, A.E.; Turnbull, D.M. Expanding Our Understanding of MtDNA Deletions. Cell Metab. 2016, 24, 3–4. [Google Scholar] [CrossRef] [Green Version]
- Bris, C.; Goudenege, D.; Desquiret-Dumas, V.; Charif, M.; Colin, E.; Bonneau, D.; Amati-Bonneau, P.; Lenaers, G.; Reynier, P.; Procaccio, V. Bioinformatics Tools and Databases to Assess the Pathogenicity of Mitochondrial DNA Variants in the Field of Next Generation Sequencing. Front. Genet. 2018, 9, 632. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.; Xie, X.; Uhler, J.P.; Hedberg-Oldfors, C.; Milenkovic, D.; Baris, O.R.; Kimoloi, S.; Matic, S.; Stewart, J.B.; Larsson, N.-G.; et al. Accurate Mapping of Mitochondrial DNA Deletions and Duplications Using Deep Sequencing. PLoS Genet. 2020, 16, e1009242. [Google Scholar] [CrossRef]
- Bris, C.; Goudenège, D.; Desquiret-Dumas, V.; Gueguen, N.; Bannwarth, S.; Gaignard, P.; Rucheton, B.; Trimouille, A.; Allouche, S.; Rouzier, C.; et al. Improved Detection of Mitochondrial DNA Instability in Mitochondrial Genome Maintenance Disorders. Genet. Med. 2021, 23, 1769–1778. [Google Scholar] [CrossRef]
- Lujan, S.A.; Longley, M.J.; Humble, M.H.; Lavender, C.A.; Burkholder, A.; Blakely, E.L.; Alston, C.L.; Gorman, G.S.; Turnbull, D.M.; McFarland, R.; et al. Ultrasensitive Deletion Detection Links Mitochondrial DNA Replication, Disease, and Aging. Genome Biol. 2020, 21, 248. [Google Scholar] [CrossRef]
- Van de Perre, P.; Goga, A.; Ngandu, N.; Nagot, N.; Moodley, D.; King, R.; Molès, J.-P.; Mosqueira, B.; Chirinda, W.; Scarlatti, G.; et al. Eliminating Postnatal HIV Transmission in High Incidence Areas: Need for Complementary Biomedical Interventions. Lancet 2021, 397, 1316–1324. [Google Scholar] [CrossRef]
- AIDSinfo|UNAIDS. Available online: http://aidsinfo.unaids.org/ (accessed on 10 December 2020).
- Afran, L.; Garcia Knight, M.; Nduati, E.; Urban, B.C.; Heyderman, R.S.; Rowland-Jones, S.L. HIV-Exposed Uninfected Children: A Growing Population with a Vulnerable Immune System? Clin. Exp. Immunol. 2014, 176, 11–22. [Google Scholar] [CrossRef]
- Wedderburn, C.J.; Evans, C.; Yeung, S.; Gibb, D.M.; Donald, K.A.; Prendergast, A.J. Growth and Neurodevelopment of HIV-Exposed Uninfected Children: A Conceptual Framework. Curr. HIV/AIDS Rep. 2019, 16, 501–513. [Google Scholar] [CrossRef] [Green Version]
- Margolis, A.M.; Heverling, H.; Pham, P.A.; Stolbach, A. A Review of the Toxicity of HIV Medications. J. Med. Toxicol. 2014, 10, 26–39. [Google Scholar] [CrossRef] [Green Version]
- Schank, M.; Zhao, J.; Moorman, J.P.; Yao, Z.Q. The Impact of HIV- and ART-Induced Mitochondrial Dysfunction in Cellular Senescence and Aging. Cells 2021, 10, 174. [Google Scholar] [CrossRef]
- Divi, R.L.; Walker, V.E.; Wade, N.A.; Nagashima, K.; Seilkop, S.K.; Adams, M.E.; Nesel, C.J.; O’neill, J.P.; Abrams, E.J.; Poirier, M.C. Mitochondrial Damage and DNA Depletion in Cord Blood and Umbilical Cord from Infants Exposed in Utero to Combivir. Aids 2004, 18, 1013–1021. [Google Scholar] [CrossRef]
- Ajaykumar, A.; Zhu, M.; Kakkar, F.; Brophy, J.; Bitnun, A.; Alimenti, A.; Soudeyns, H.; Saberi, S.; Albert, A.Y.K.; Money, D.M.; et al. Blood Mitochondrial DNA Levels Remain Elevated from Birth to Early Life in Children HIV-Exposed Uninfected Exposed to Combination Antiretroviral Therapy in Utero. J. Infect. Dis. 2020, 223, 621–629. [Google Scholar] [CrossRef]
- Monnin, A.; Nagot, N.; Periès, M.; Vallo, R.; Meda, N.; Singata-Madliki, M.; Tumwine, J.K.; Kankasa, C.; Ngandu, N.; Goga, A.; et al. Mitochondrial DNA Parameters in Blood of Infants Receiving Lopinavir/Ritonavir or Lamivudine Prophylaxis to Prevent Breastfeeding Transmission of HIV-1. J. Clin. Med. 2020, 9, E2972. [Google Scholar] [CrossRef]
- Ouyang, Y.; Qiao, L.; Liu, K.; Zang, Y.; Sun, Y.; Dong, Y.; Liu, D.; Guo, X.; Wei, F.; Lin, M.; et al. Mitochondrial DNA Mutations in Blood Samples from HIV-1-Infected Children Undergoing Long-Term Antiretroviral Therapy. Mutat. Res. Genet. Toxicol. Env. Mutagen 2016, 805, 1–6. [Google Scholar] [CrossRef]
- Ouyang, Y.; Wei, F.; Qiao, L.; Liu, K.; Dong, Y.; Guo, X.; Wang, S.; Pang, L.; Lin, M.; Zhang, F.; et al. Mitochondrial DNA Mutations Accumulated in HIV-1-Infected Children Who Have an Excellent Virological Response When Exposed to Long-Term Antiretroviral Therapy. J. Antimicrob. Chemother. 2018, 73, 3114–3121. [Google Scholar] [CrossRef]
- Roca-Bayerri, C.; Robertson, F.; Pyle, A.; Hudson, G.; Payne, B.A.I. Mitochondrial DNA Damage and Brain Aging in Human Immunodeficiency Virus. Clin. Infect. Dis. 2021, 73, e466–e473. [Google Scholar] [CrossRef]
- Roda, R.H.; Bargiela, D.; Chen, W.; Perry, K.; Ellis, R.J.; Clifford, D.B.; Bharti, A.; Kallianpur, A.R.; Oliveira, M.F.; Diaz, M.M.; et al. Large Mitochondrial DNA Deletions in HIV Sensory Neuropathy. Neurology 2021, 97, e156–e165. [Google Scholar] [CrossRef]
- Solanky, D.; Fields, J.A.; Iudicello, J.E.; Ellis, R.J.; Franklin, D.; Clifford, D.B.; Gelman, B.B.; Marra, C.M.; Morgello, S.; Rubin, L.H.; et al. Higher Buccal Mitochondrial DNA and Mitochondrial Common Deletion Number Are Associated with Markers of Neurodegeneration and Inflammation in Cerebrospinal Fluid. J. Neurovirol. 2022, 28, 281–290. [Google Scholar] [CrossRef]
- Ziada, A.S.; Lu, M.Y.; Ignas-Menzies, J.; Paintsil, E.; Li, M.; Ogbuagu, O.; Saberi, S.; Hsieh, A.Y.Y.; Sattha, B.; Harrigan, P.R.; et al. Mitochondrial DNA Somatic Mutation Burden and Heteroplasmy Are Associated with Chronological Age, Smoking, and HIV Infection. Aging Cell 2019, 18, e13018. [Google Scholar] [CrossRef] [Green Version]
- Monnin, A.; Desquiret-Dumas, V.; Méda, N.; Goudenège, D.; Bris, C.; Kankasa, C.; Singata-Madliki, M.; Tylleskar, T.; Procaccio, V.; Nagot, N.; et al. Mitochondrial DNA Instability Is Common in HIV-Exposed Uninfected Newborns. J. Clin. Med. 2021, 10, 2399. [Google Scholar] [CrossRef]
- Monnin, A.; Nagot, N.; Eymard-Duvernay, S.; Meda, N.; Tumwine, J.K.; Tylleskär, T.; de Perre, P.V.; Molès, J.-P. Health Outcomes at School Age among Children Who Are HIV-Exposed but Uninfected with Detected Mitochondrial DNA Depletion at One Year. J. Clin. Med. 2020, 9, E3680. [Google Scholar] [CrossRef]
- Nagot, N.; Kankasa, C.; Tumwine, J.K.; Meda, N.; Hofmeyr, G.J.; Vallo, R.; Mwiya, M.; Kwagala, M.; Traore, H.; Sunday, A.; et al. Extended Pre-Exposure Prophylaxis with Lopinavir-Ritonavir versus Lamivudine to Prevent HIV-1 Transmission through Breastfeeding up to 50 Weeks in Infants in Africa (ANRS 12174): A Randomised Controlled Trial. Lancet 2016, 387, 566–573. [Google Scholar] [CrossRef]
- Nagot, N.; Singata-Madliki, M.; Cournil, A.; Nalugya, J.; Tassembedo, S.; Quillet, C.; Tonga, M.W.; Tumwine, J.; Meda, N.; Kankasa, C.; et al. Growth, Clinical and Neurodevelopmental Outcomes at School Age Are Similar for Children Who Received 1-Year Lamivudine or Lopinavir/Ritonavir HIV Prophylaxis in Early Life. Sci. Rep. 2021, 4, 3173. [Google Scholar] [CrossRef]
- WHO. Child Growth Standards: Length/Height-for-Age, Weight-for-Age, Weight-for-Length, Weight-for-Height and Body Mass Index-for-Age: Methods and Development. Available online: https://www.who.int/publications-detail-redirect/924154693X (accessed on 11 April 2022).
- Calculators: Blood Pressure Percentiles for Boys (2–17 years). Available online: https://www.msdmanuals.com/professional/multimedia/clinical-calculator/blood-pressure-percentiles-for-boys-2-17-years (accessed on 11 April 2022).
- DAIDS. Adverse Event Grading Tables|DAIDS Regulatory Support Center (RSC). Available online: https://rsc.niaid.nih.gov/clinical-research-sites/daids-adverse-event-grading-tables (accessed on 12 April 2022).
- Goudenège, D.; Bris, C.; Hoffmann, V.; Desquiret-Dumas, V.; Jardel, C.; Rucheton, B.; Bannwarth, S.; Paquis-Flucklinger, V.; Lebre, A.S.; Colin, E.; et al. EKLIPse: A Sensitive Tool for the Detection and Quantification of Mitochondrial DNA Deletions from next-Generation Sequencing Data. Genet. Med. 2019, 21, 1407–1416. [Google Scholar] [CrossRef]
- Jang, J.Y.; Blum, A.; Liu, J.; Finkel, T. The Role of Mitochondria in Aging. J. Clin. Investig. 2018, 128, 3662–3670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Contreras, M.; Kennedy, S.R. The Complicated Nature of Somatic MtDNA Mutations in Aging. Front. Aging 2022, 2, 805126. [Google Scholar] [CrossRef] [PubMed]
- Blackwood, J.K.; Whittaker, R.G.; Blakely, E.L.; Alston, C.L.; Turnbull, D.M.; Taylor, R.W. The Investigation and Diagnosis of Pathogenic Mitochondrial DNA Mutations in Human Urothelial Cells. Biochem. Biophys. Res. Commun. 2010, 393, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Lawless, C.; Greaves, L.; Reeve, A.K.; Turnbull, D.M.; Vincent, A.E. The rise and rise of mitochondrial DNA mutations. Open Biol. 2020, 10, 200061. [Google Scholar] [CrossRef]
- Fontana, G.A.; Gahlon, H.L. Mechanisms of Replication and Repair in Mitochondrial DNA Deletion Formation. Nucleic Acids Res. 2020, 48, 11244–11258. [Google Scholar] [CrossRef]
- Gutiérrez-Sevilla, J.E.; Cárdenas-Bedoya, J.; Escoto-Delgadillo, M.; Zúñiga-González, G.M.; Pérez-Ríos, A.M.; Gómez-Meda, B.C.; González-Enríquez, G.V.; Figarola-Centurión, I.; Chavarría-Avila, E.; Torres-Mendoza, B.M. Genomic Instability in People Living with HIV. Mutat. Res. Genet. Toxicol. Env. Mutagen 2021, 865, 503336. [Google Scholar] [CrossRef]
- Grando, A.C.; Guimarães, N.N.; de Souza, A.P.; Lehmann, M.; Cunha, K.S.; Dihl, R.R. Assessment of Complex Genomic Alterations Induced by AZT, 3TC, and the Combination AZT +3TC. Drug Chem. Toxicol. 2020, 43, 429–434. [Google Scholar] [CrossRef]
- Teodorof-Diedrich, C.; Spector, S.A. Human Immunodeficiency Virus Type 1 gp120 and Tat Induce Mitochondrial Fragmentation and Incomplete Mitophagy in Human Neurons. J. Virol. 2018, 92, e00993-18. [Google Scholar] [CrossRef] [Green Version]
- Divi, R.L.; Einem, T.L.; Fletcher, S.L.L.; Shockley, M.E.; Kuo, M.M.; St Claire, M.C.; Cook, A.; Nagashima, K.; Harbaugh, S.W.; Harbaugh, J.W.; et al. Progressive Mitochondrial Compromise in Brains and Livers of Primates Exposed in Utero to Nucleoside Reverse Transcriptase Inhibitors (NRTIs). Toxicol. Sci. 2010, 118, 191–201. [Google Scholar] [CrossRef]
- Liu, Y.; Shim Park, E.; Gibbons, A.T.; Shide, E.D.; Divi, R.L.; Woodward, R.A.; Poirier, M.C. Mitochondrial Compromise in 3-Year Old Patas Monkeys Exposed in Utero to Human-Equivalent Antiretroviral Therapies. Env. Mol. Mutagen 2016, 57, 526–534. [Google Scholar] [CrossRef] [Green Version]
- Bañó, M.; Morén, C.; Barroso, S.; Juárez, D.L.; Guitart-Mampel, M.; González-Casacuberta, I.; Canto-Santos, J.; Lozano, E.; León, A.; Pedrol, E.; et al. Mitochondrial Toxicogenomics for Antiretroviral Management: HIV Post-Exposure Prophylaxis in Uninfected Patients. Front. Genet. 2020, 11, 497. [Google Scholar] [CrossRef]
- Republic of Zambia; Ministry of Health Zambia. Consolidated Guidelines for Treatment and Prevention of HIV Infection; Ministry of Health Zambia: Lusaka, Zambia, 2020. [Google Scholar]
- Stoker, M.L.; Newport, E.; Hulit, J.C.; West, A.P.; Morten, K.J. Impact of Pharmacological Agents on Mitochondrial Function: A Growing Opportunity? Biochem. Soc. Trans. 2019, 47, 1757–1772. [Google Scholar] [CrossRef] [Green Version]
- Pitceathly, R.D.S.; Keshavan, N.; Rahman, J.; Rahman, S. Moving towards Clinical Trials for Mitochondrial Diseases. J. Inherit. Metab. Dis. 2021, 44, 22–41. [Google Scholar] [CrossRef]



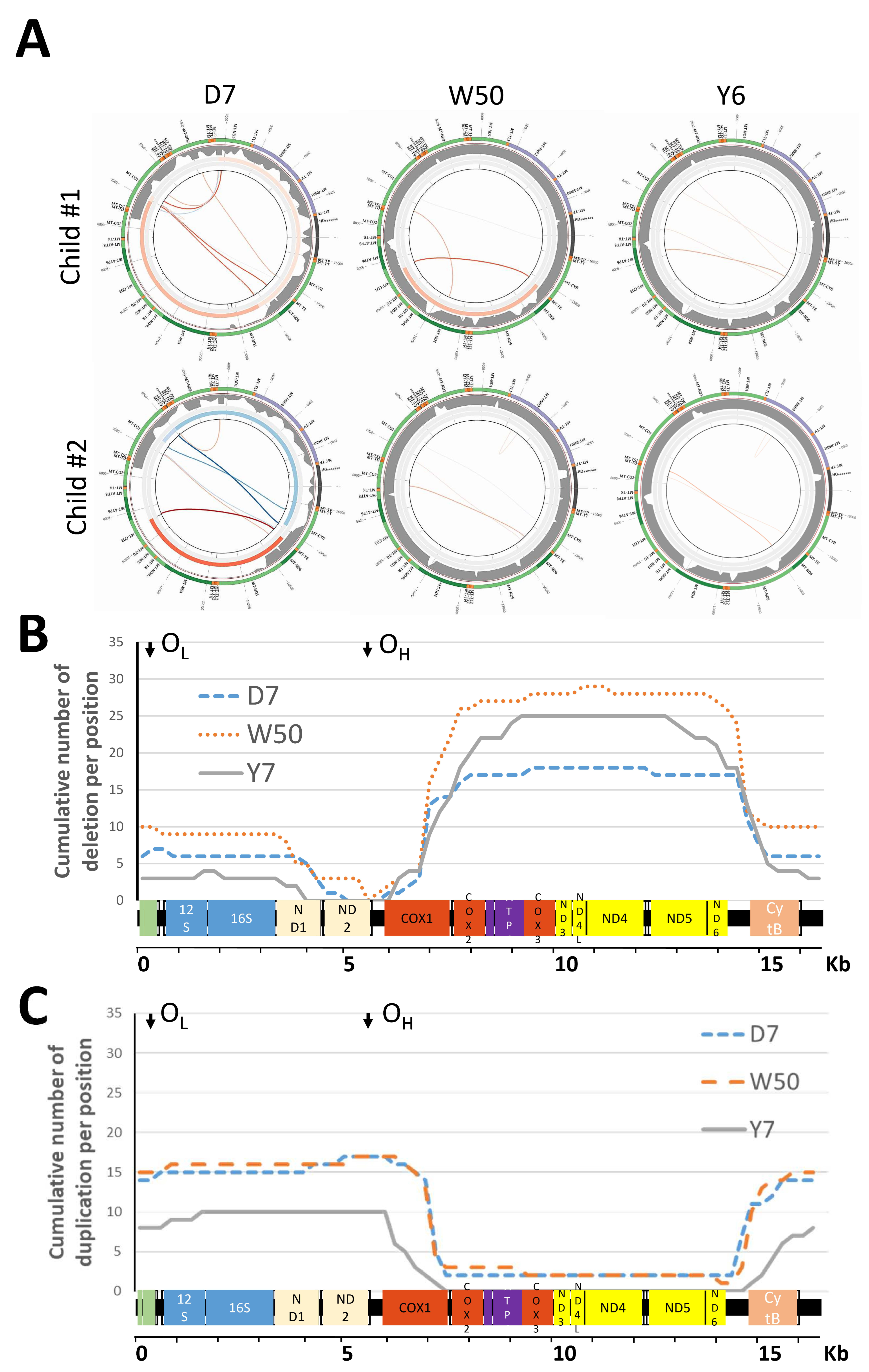{kind=link}
{kind=link}
| D7 | W50 | Y6 | |
|---|---|---|---|
| Socio-Demographics | |||
| Site; n (%) | |||
| Burkina Faso | 13 (54.2) | ||
| Zambia | 11 (45.8) | ||
| Sex; n (%) | |||
| Boy | 12 (50.0) | ||
| Age (in weeks/years); mean ± SD | 1.0 ± 0.0 | 1.0 ± 0.1 † | 6.3 ± 0.3 |
| Anthropometrics; mean ± SD | |||
| Weight (kg) | 3.1 ± 0.4 | 8.1 ± 1.1 † | 19.1 ± 2.9 |
| Height (cm) | 49.0 ± 2.2 | 72.3 ± 3.0 † | 114.1 ± 5.2 |
| Head circumference (cm) | N.A. | N.A. | 50.8 ± 1.7 † |
| WAZ | −0.7 ± 0.9 | −1.2 ± 1.1 † | −0.8 ± 1.0 |
| HAZ | −0.9 ± 1.1 | −0.9 ± 1.2 † | −0.6 ± 1.0 |
| BMIZ | −0.4 ± 1.5 † | −0.9 ± 1.1 † | −0.6 ± 0.9 |
| Gestational age (week); median [IQR] | 38.0 [37.0;39.5] | N.A. | N.A. |
| Preterm birth (week); n (%) | |||
| No prematurity ≥ 37 | 21 (87.5) | ||
| Prematurity < 37 | 3 (12.5) | ||
| General Examination; n (%) | |||
| Blood pressure, systolic (mmHg) | |||
| Low < 95 | 12 (50.0) | ||
| Normal (95–100) | N.A. | N.A. | 3 (12.5) |
| Elevated > 100 | 9 (37.5) | ||
| Elevated blood pressure, diastolic (mmHg) | |||
| Normal (56–70) | N.A. | N.A. | 14 (58.3) |
| Elavated > 70 | 10 (41.7) | ||
| Elevated pulse rate (rate/min) | |||
| Normal (75–110) | N.A. | N.A. | 23 (95.8) |
| Elevated > 110 | 1 (4.2) | ||
| Elevated respiratory rate (rate/min) | |||
| Normal (21–23) | N.A. | N.A. | 9 (37.5) |
| Elevated > 23 | 15 (62.5) | ||
| Oedema; n (%) | 0 (0.0) | 0 (0.0) | |
| Clinical evaluation | |||
| No abnormality | 20 (83.3) | ||
| One abnormality | N.A. | N.A. | 4 (16.7) |
| Blindness | 1 (4.2) | ||
| Hearing impairment | 1 (4.2) | ||
| Mild Arrhythmia | 1 (4.2) | ||
| Heart murmur | 1 (4.2) | ||
| Hematology; n (%) | |||
| Hemoglobin concentration (g/dL); n (%) | |||
| Normal | 22 (91.7) | 9 (52.9) §,° | 23 (95.8) |
| Anemia # | 2 (8.3) | 8 (47.1) §,° | 1 (4.2) |
| Platelet count (103/mm3); n (%) | |||
| Normal > 125 | 24 (100.0) | 17 (100.0) §,° | 24 (100.0) |
| Leucocyte count (103/mm3); n (%) | |||
| Normal > 2.5 | 24 (100.0) | 17 (100.0) §,° | 24 (100.0) |
| Neutrophil count (103/mm3); n (%) | |||
| Normal | 23 (100.0) † | 14 (82.4) §,° | 24 (100.0) |
| Neutropenia ¥ | 0.0 (0.0) | 6 (17.7) §,° | |
| Biochemistry; n (%) | |||
| LDH concentration (xULN) | N.A. | N.A. | |
| Normal < ULN | N.A. | 7 (29.2) | |
| Abnormal ≥ ULN | N.A. | 17 (70.8) | |
| Grade 1 (Mild) [ULN-2xULN] | N.A. | 12 (50.0) | |
| Grade 2 (Moderate) [≥2xULN] | N.A. | 5 (20.8) | |
| ALT concentration (U/L) | |||
| Normal < 1.25xULN | 24 (100.0) | 16 (94.1) §,° | 24 (100.0) |
| Abnormal ≥ 1.25xULN | 1 (5.9) §,° | ||
| Grade 1 (Mild) [1.25–2.5]xULN | 1 (5.9) §,° | ||
| Medical events; n (%) | |||
| Clinical consultation without admission during the last year | |||
| Yes | N.A. | N.A. | 19 (79.2) |
| Hospital admission since the PROMISE-PEP trial | |||
| Yes | N.A. | N.A. | 5 (20.8) |
| Child ARV prophylaxis during the PROMISE-PEP trial | |||
| Lopinavir/ritonavir (LPV/r) | 12 (50.0) | 12 (50.0) | |
| Development and neuropsychological assessment | |||
| 10questionsPlus | |||
| At least 1 disability; n (%) | N.A. | N.A. | 7 (29.2) |
| SDQ-25; mean ± SD | N.A. | N.A. | 7.7 ± 4.5 |
| TOVA; mean ± SD | N.A. | N.A. | 2.2 ± 0.7 † |
| MABC-2; mean ± SD | N.A. | N.A. | 75.6 ± 9.6 ‡ |
| KABC-II; mean ± SD | N.A. | N.A. | 57.4 ± 13.8 ‡ |
| Sociodemographic characteristics | |
| Age (year); mean ± SD | 28.2 ± 5.6 |
| Parity; median [IQR] | 3.0 [1.5;3.0] |
| Education | |
| Mother/caregiver ever attended school; n (%) | |
| Yes | 18 (75.0) |
| Clinical and biological characteristics | |
| BMI; mean ± SD | 23.3 ± 2.8 |
| CD4 cells count (cells/mm3); median [IQR] | 543 [450;820] |
| HIV viral load control (copies/mL); n (%) | |
| <1000 | 14 (58.3) |
| ≥1000 | 10 (41.7) |
| WHO HIV staging; n (%) | |
| Stage 1 | 24 (100.0) |
| Mode of delivery; n (%) | |
| Vaginal | 24 (100.0) |
| Maternal prophylaxis during pregnancy | |
| ARV regimen; n (%) | |
| AZT | 24 (100.0) |
| Duration of ARV prophylaxis (week); median [IQR] | 8.0 [4.0;9.0] |
| Maternal lifestyle during pregnancy | |
| Smoking during pregnancy; n (%) | |
| No | 22 (100.0) ‡ |
| Alcohol consumption during pregnancy; n (%) | |
| Yes No | 12 (54.6) ‡ 10 (45.5) ‡ |
| D7 (n = 24) | W50 (n = 24) | Y6 (n = 24) | |
|---|---|---|---|
| Child with mtDNA alteration; n (%) | 16 (66.7) | 20 (83.3) | 18 (75.0) |
| Alteration events; n | 36 | 50 | 33 |
| Nb of event per affected child | 2 [1.0–2.5] | 2.0 [1.5–3.0] | 1.0 [1.0–3.0] |
| Deletion | 20 | 33 | 23 |
| Class I/II/III | 4/10/8 | 4/11/18 | 5/6/12 |
| Duplication | 16 | 17 | 10 |
| Cumulative heteroplasmy (in %) | |||
| Median [IQR] | 0.24 [0.00–2.47] | 0.43 [0.19–2.46] | 0.24 [0.05–0.64] |
| Deletion heteroplasmy (in %) | |||
| Median [IQR] | 0.00 [0.00–0.48] | 0.40 [0.06–1.38] | 0.16 [0.00–0.36] |
| Duplication heteroplasmy (in %) | |||
| Median [IQR] | 0.03 [0.00–0.15] | 0.00 [0.00–0.13] | 0.00 [0.00–0.11] |
| Child with heteroplasmy > 1%; n (%) | 7 (29.2) | 9 (37.5) | 3 (12.5) |
| Point mutation; n | 0 | 0 | 0 |
| Child receiving 3TC PrEP (n = 12) | |||
| Child with mtDNA alteration; n (%) | 9 (75.0) | 10 (83.3) | 8 (66.7) |
| Alteration events; n | 22 | 26 | 12 |
| Cumulative heteroplasmy | |||
| Median [IQR] | 0.41 [0.07–3.68] | 0.32 [0.15–2.46] | 0.13 [0.00–0.48] |
| Deletion class I/II/III | 3/6/4 | 1/4/10 | 1/2/5 |
| Child receiving LPV/r PrEP (n = 12) | |||
| Child with mtDNA alteration; n (%) | 7 (58.3) | 10 (83.3) | 10 (83.3) |
| Alteration events; n | 14 | 24 | 20 |
| Cumulative heteroplasmy | |||
| Median [IQR] | 0.08 [0.00–0.76] | 0.54 [0.23–2.46] | 0.35 [0.18–0.71] |
| Deletion class I/II/III | 1/4/7 | 3/7/8 | 4/4/10 |
| N | Cumulative Hetereoplasmy Rate at D7 | p Value | ||
|---|---|---|---|---|
| n (%) > 1% | ρ | |||
| Site | ||||
| Burkina Faso | 13 | 5 (38.5) | 0.36 | |
| Zambia | 11 | 2 (18.2) | ||
| Sex | ||||
| Boy | 12 | 4 (33.3) | 1.000 | |
| Girl | 12 | 3 (25.0) | ||
| Preterm birth | ||||
| No prematurity (≥37 weeks) | 21 | 3 (33.3) | 0.530 | |
| Premature (<37 weeks) | 3 | 0 (0.0) | ||
| Parity (per child) | 24 | 0.034 | 0.876 | |
| Child weight at D7 (per kg) | 24 | −0.257 | 0.225 | |
| Mother’s age at birth | ||||
| ≤30 years | 14 | 4 (28.6) | 0.643 | |
| 30 years and over | 10 | 3 (30.0) | ||
| Mother education | ||||
| Yes | 18 | 4 (22.2) | 0.307 | |
| No | 6 | 3 (50.0) | ||
| Mother’s HIV viral load | ||||
| <1000 copies/mL | 14 | 4 (28.6) | 1.000 | |
| ≥1000 copies/mL | 10 | 3 (30.0) | ||
| Duration of ARV prophylaxis during pregnancy (per week) | 24 | 0.243 | 0.253 | |
| Alcohol consumption during pregnancy | ||||
| No | 10 | 3 (30.0) | 0.624 | |
| Yes | 12 | 2 (16.7) | ||
| Health Outcomes at Y6 | Cumulative Heteroplasmy | p-Value | |
|---|---|---|---|
| rate > 1% at D7 n = 7 | rate ≤ 1% at D7 n = 17 | ||
| Growthmean±SD | |||
| Head circumference (cm) | 50.7 ± 1.7 | 50.9 ± 1.7 † | 0.777 |
| WAZ | −1.2 ± 0.7 | −0.6 ± 1.1 | 0.142 |
| HAZ | −0.9 ± 0.9 | −0.4 ± 1.1 | 0.356 |
| BMIZ | −1.0 ± 0.9 | −0.4 ± 0.9 | 0.159 |
| Clinical evaluation | |||
| Systolic blood pressure Z-score; mean ± SD | 0.7 ± 1.0 | 0.3 ± 1.1 | 0.463 |
| Clinical abnormalities; n (%) | 1 (14.3) | 3 (17.7) | 1.000 |
| Hospitalised since PROMISE PEP; n (%) | 3 (42.9) | 2 (11.8) | 0.126 |
| Abnormal LDH concentration; n (%) | 5 (71.4) | 12 (70.6) | 1.000 |
| Abnormal ALT concentration; n (%) | 0 (0.0) | 0 (0.0) | N.A. |
| Developmental outcomes | |||
| At least one disability detected from the 10questionPlus; n (%) | 3 (42.9) | 4 (23.5) | 0.374 |
| SDQ-25; mean ± SD | 9.1 ± 4.4 | 7.1 ± 4.5 | 0.326 |
| TOVA; mean ± SD | 2.1 ± 0.3 | 2.3 ± 0.8 † | 0.622 |
| MABC-2; mean ± SD | 74.8 ± 9.0 † | 75.9 ± 10.1 † | 0.827 |
| KABC-II; mean ± SD | 54.4 ± 21.5 † | 58.5 ± 10.4 † | 0.558 |
| Health Outcomes at Y6 | β [95%CI] for Duration of Maternal Prophylaxis, Stratified by Infant Heteroplasmy Rate | p-Value for Interaction | ||
|---|---|---|---|---|
| Total N = 24 | >1% at D7 n = 7 | ≤1% at D7 n = 17 | ||
| Growth | ||||
| Head circumference (cm) † | 0.20 [−0.05–0.44] | 0.31 [0.06–0.55] | −0.40 [−1.19–0.38] | 0.033 * |
| WAZ | 0.02 [−0.13–0.18] | −0.13 [−0.46–0.20] | 0.06 [−0.10–0.23] | 0.364 |
| HAZ | 0.06 [−0.10–0.22] | 0.14 [−0.32–0.61] | 0.05 [−0.12–0.23] | 0.685 |
| BMIZ | −0.02 [−0.17–0.12] | −0.34 [−0.66–−0.02] | 0.04 [−0.10–0.19] | 0.047 * |
| Clinical evaluation | ||||
| Systolic BP Z-score | 0.09 [−0.06–0.24] | 0.15 [−0.33–0.63] | 0.08 [−0.10–0.25] | 0.733 |
| LDH concentration | −6.79 [−68.63–55.05] | 20.91 [−65.57–107.39] | −7.55 [−76.26–61.16] | 0.255 |
| ALT concentration | 0.40 [−0.76–1.56] | 2.87 [−2.30–8.05] | −0.05 [−1.27–1.16] | 0.070 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Desquiret-Dumas, V.; D’Ottavi, M.; Monnin, A.; Goudenège, D.; Méda, N.; Vizeneux, A.; Kankasa, C.; Tylleskar, T.; Bris, C.; Procaccio, V.; et al. Long-Term Persistence of Mitochondrial DNA Instability in HIV-Exposed Uninfected Children during and after Exposure to Antiretroviral Drugs and HIV. Biomedicines 2022, 10, 1786. https://doi.org/10.3390/biomedicines10081786
Desquiret-Dumas V, D’Ottavi M, Monnin A, Goudenège D, Méda N, Vizeneux A, Kankasa C, Tylleskar T, Bris C, Procaccio V, et al. Long-Term Persistence of Mitochondrial DNA Instability in HIV-Exposed Uninfected Children during and after Exposure to Antiretroviral Drugs and HIV. Biomedicines. 2022; 10(8):1786. https://doi.org/10.3390/biomedicines10081786
Chicago/Turabian StyleDesquiret-Dumas, Valérie, Morgana D’Ottavi, Audrey Monnin, David Goudenège, Nicolas Méda, Amélie Vizeneux, Chipepo Kankasa, Thorkild Tylleskar, Céline Bris, Vincent Procaccio, and et al. 2022. "Long-Term Persistence of Mitochondrial DNA Instability in HIV-Exposed Uninfected Children during and after Exposure to Antiretroviral Drugs and HIV" Biomedicines 10, no. 8: 1786. https://doi.org/10.3390/biomedicines10081786