
SlHSP17.7 Ameliorates Chilling Stress-Induced Damage by Regulating Phosphatidylglycerol Metabolism and Calcium Signal in Tomato Plants


Abstract
:1. Introduction
2. Results
2.1. SlHSP17.7 Overexpression Improved Chilling Tolerance of Tomato Plants
2.2. SlHSP17.7 Overexpression Improved Photosynthesis under Chilling Stress
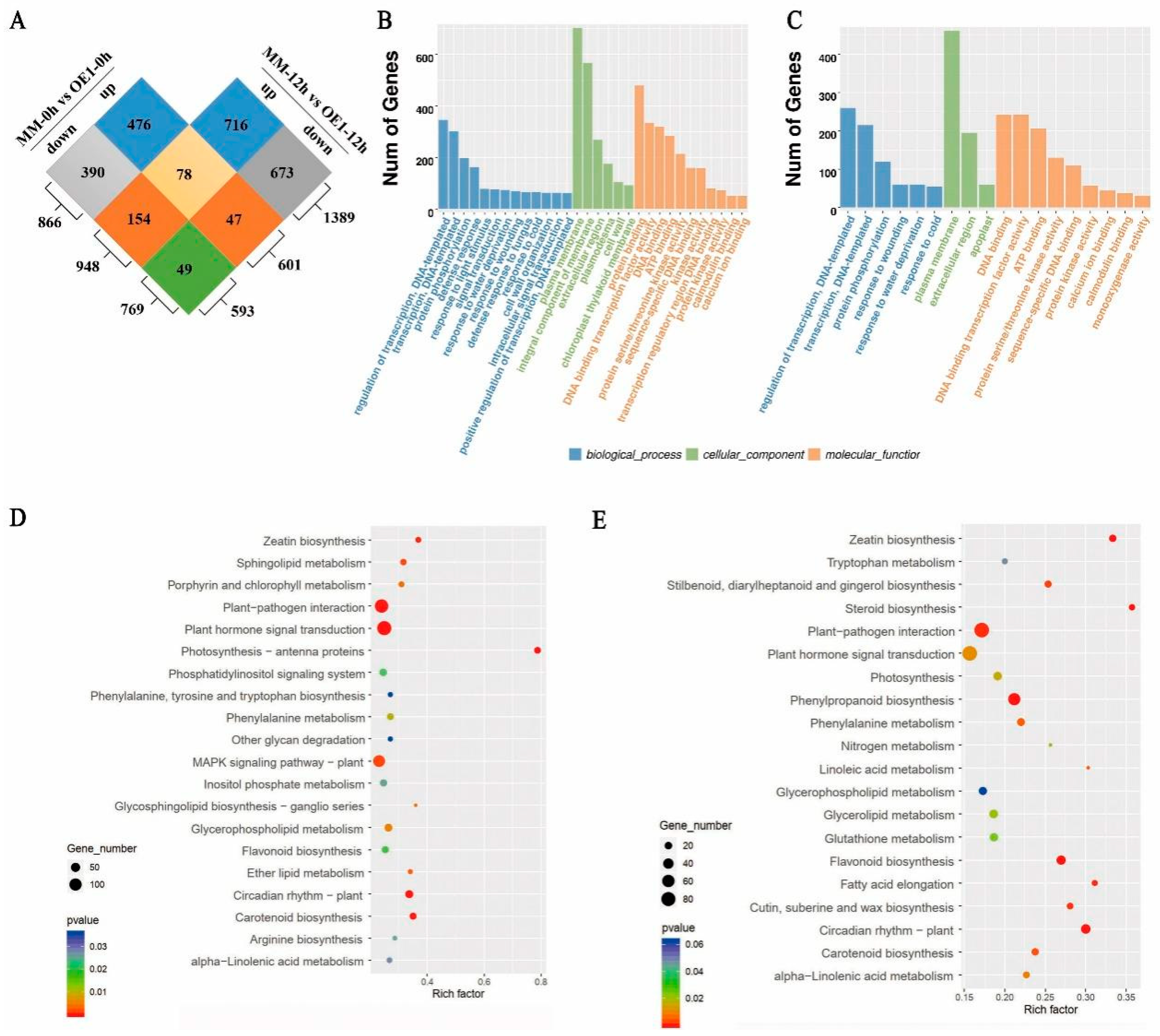
2.3. SLHSP17.7 Overexpression Initiated a Large Transcriptome Reprogramming under Chilling Stress
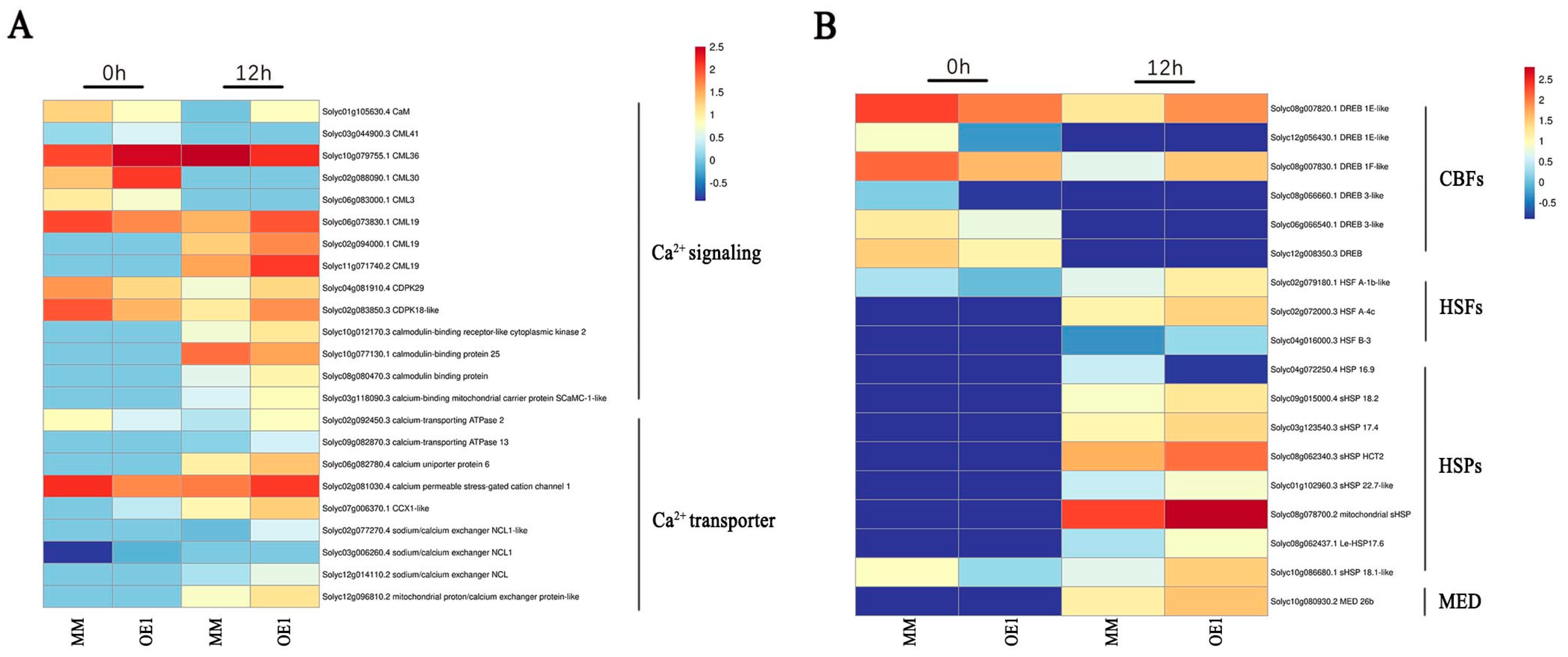
2.4. SlHSP17.7 Overexpression Activated Ca2+ Signaling and CBF Transcription Factors under Chilling Stress
2.5. SlHSP17.7 Overexpression Positively Regulated Reactive Oxygen Species (ROS)-Scavenging-Related Genes and Alleviated ROS Accumulation under Chilling Stress
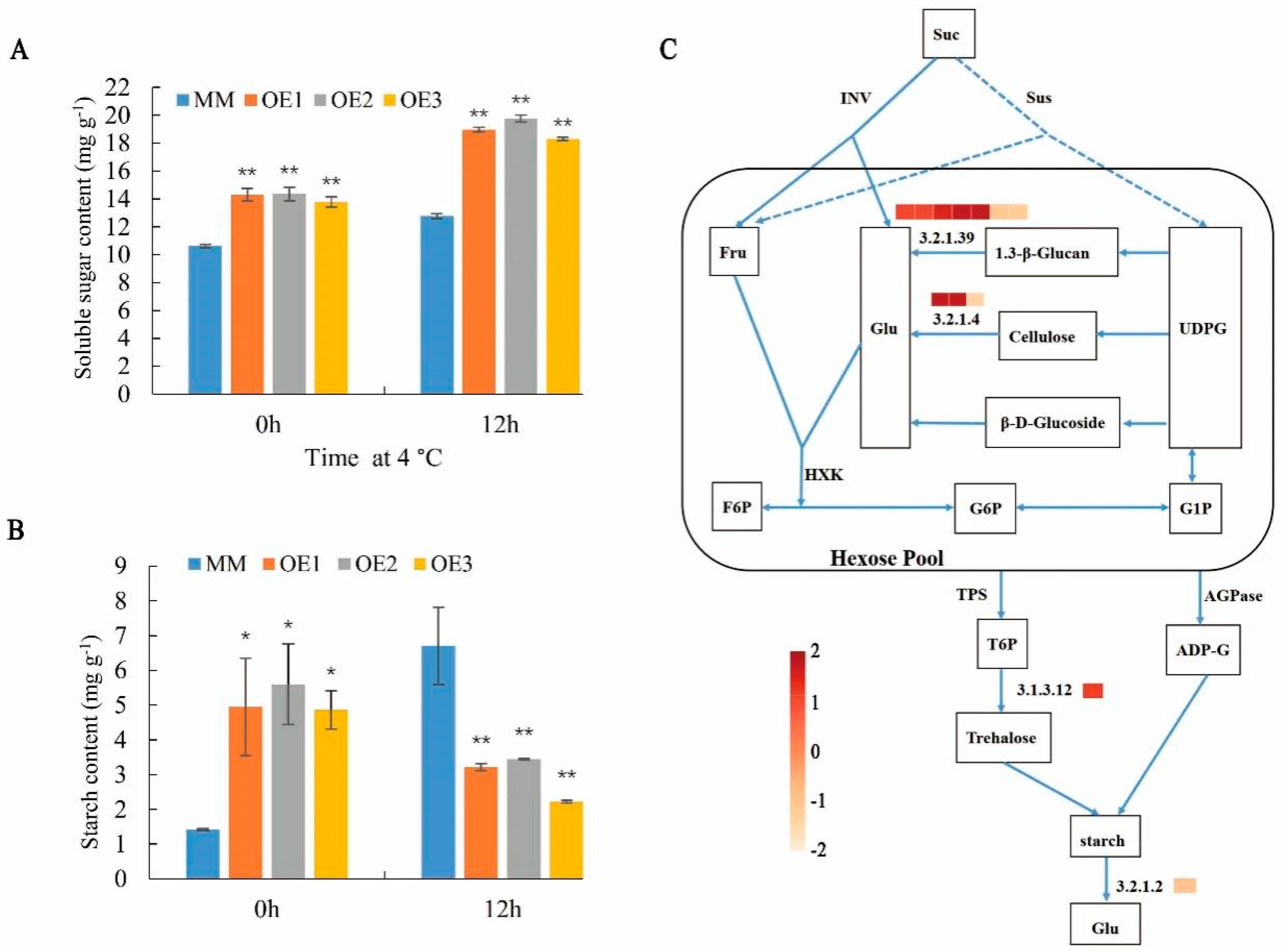
2.6. SlHSP17.7 Overexpression Promoted Starch Formation under Normal Conditions and Accumulates More Soluble Sugars under Chilling Stress
2.7. SlHSP17.7 Overexpression Improved the Fluidity of Cell Membrane and Phosphatidylglycerol (PG) Metabolism upon Metabolite Identification
3. Discussion
4. Materials and Methods
4.1. Materials and Chilling Stress Treatments
4.2. Evaluation of Chilling Injury Index
4.3. Measurement of Physiological Parameters
4.4. Observation of Chloroplast Ultrastructure
4.5. RNA Sequencing Analysis
4.6. qRT-PCR Analysis
4.7. Cloning MED26b and VIGS Vector Construction
4.8. Agrobacterium Preparation and Inoculation
4.9. Metabolite Extraction and Analysis
4.10. Data Analysis and Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thakur, P.; Nayyar, H. Facing the cold stress by plants in the changing environment: Sensing, signaling, and defending mechanisms. In Plant Acclimation to Environmental Stress; Springer: New York, NY, USA, 2012; Volume 2, pp. 29–69. [Google Scholar] [CrossRef]
- Barrero-Gil, J.; Huertas, R.; Rambla, J.L.; Granell, A.; Salinas, J. Tomato plants increase their tolerance to low temperature in a chilling acclimation process entailing comprehensive transcriptional and metabolic adjustments. Plant Cell Environ. 2016, 39, 2303–2318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyons, J.M. Chilling injury in plants. Annu. Rev. Plant Physiol. 1973, 24, 445–466. [Google Scholar] [CrossRef]
- Zheng, D.H.; Huang, J.X.; Wang, L.J. Research progress in the ecology of tomato at low temperature. North. Hortic. 2002, 3, 36–37. [Google Scholar]
- Wang, N.; Fang, W.; Han, H.; Sui, N.; Li, B.; Meng, Q.W. Overexpression of zeaxanthin epoxidase gene enhances the sensitivity of tomato PSII photoinhibition to high light and chilling stress. Physiol. Plantarum. 2008, 132, 384–396. [Google Scholar] [CrossRef]
- Zhang, X.; Fowler, S.G.; Cheng, H.; Lou, Y.; Rhee, S.Y.; Stockinger, E.J.; Thomashow, M.F. Freezing-sensitive tomato has a functional CBF cold response pathway, but a CBF regulon that differs from that of freezing tolerant Arabidopsis. Plant J. 2004, 39, 905–919. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Guo, Z.; Li, H.; Wang, M.; Onac, E.; Zhou, J.; Xia, X.J.; Shi, K.; Yu, J.Q.; Zhou, Y.H. Phytochrome A and B function antagonistically to regulate cold tolerance via abscisic acid-dependent jasmonate signaling. Plant Physiol. 2016, 170, 459–471. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.Y.; Jiang, X.C.; Liu, Q.Y.; Ahammed, G.J.; Lin, R.; Wang, L.Y.; Shao, S.J.; Yu, J.Q.; Zhou, Y.H. The HY5 and MYB15 transcription factors positively regulate cold tolerance in tomato via the CBF pathway. Plant Cell Environ. 2020, 43, 2712–2726. [Google Scholar] [CrossRef] [PubMed]
- Waters, E.R. The evolution, function, structure, and expression of the plant sHSPs. J. Exp. Bot. 2013, 64, 391–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.N.; Wang, B.; Sun, D.M.; Deng, X. Molecular cloning and differential expression of sHSP gene family members from the resurrection plant Boea hygrometrica in response to abiotic stresses. Biologia 2013, 68, 651–661. [Google Scholar] [CrossRef]
- Sun, W.N.; Montagu, M.V.; Verbruggen, N. Small heat shock proteins and stress tolerance in plants. Biochim. Biophys Acta—Gene Struct. Expr. 2002, 1577, 1–9. [Google Scholar] [CrossRef]
- Upadhyay, R.K.; Tucker, M.L.; Mattoo, A.K. Ethylene and ripening inhibitor modulate expression of SlHSP17.7A, B Class I small heat shock protein genes during tomato fruit ripening. Front. Plant Sci. 2020, 11, 975. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhao, H.Y.; Shi, J.W.; Wu, Y.Y.; Li, T.L.; Jing, J. Functional characterization of the small heat shock protein 17.7 gene conferring cold stress in tomato. Plant Sci. 2020, 298, 1–12. [Google Scholar] [CrossRef]
- Chauhan, H.; Khurana, N.; Nijhavan, A.; Jitendra, P.K.; Paramjit, K. The wheat chloroplastic small heat shock protein (sHSP26) is involved in seed maturation and germination and imparts tolerance to heat stress. Plant Cell Environ. 2012, 35, 1912–1931. [Google Scholar] [CrossRef]
- Zhou, Y.L.; Chen, H.H.; Chu, P.; Li, Y.; Tan, B.; Ding, Y.; Tsang, E.W.T.; Jiang, L.W.; Wu, K.Q.; Huang, S.Z. NnHSP17.5, a cytosolic class II small heat shock protein gene from Nelumbo nucifera, contributes to seed germination vigor and seedling thermotolerance in transgenic Arabidopsis. Plant Cell Rep. 2012, 31, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Park, S.M.; Kim, K.P.; Joe, M.K.; Lee, M.O.; Koo, H.J.; Hong, C.B. Tobacco class I cytosolic small heat shock proteins are under transcriptional and translational regulations in expression and heterocomplex prevails under the high temperature stress condition in vitro. Plant Cell Environ. 2015, 38, 767–776. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, W.J.; Gao, Y.K.; Pan, H.T.; Zhang, Q.X. A cytosolic class II small heat shock protein, PfHSP17.2, confers resistance to heat, cold, and salt stresses in transgenic Arabidopsis. Genet. Mol. Biol. 2018, 41, 649–660. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, C.M.; Wang, Y.J.; Liu, J. Overexpression of chloroplast-localized small molecular heat-shock protein enhances chilling tolerance in tomato plant. J. Plant Physiol. Mol. Biol. 2005, 31, 167–174. [Google Scholar]
- Guo, P.; Sui, N.; Yu, C.; Guo, S.J.; Dong, X.C.; Meng, Q.W. CaHSP18 of sweet pepper enhanced chilling tolerance of transgenic tomato plants. Plant Physiol. Commun. 2008, 44, 409–412. [Google Scholar]
- Guo, S.J.; Zhou, H.Y.; Zhang, X.S.; Li, X.G.; Meng, Q.W. Overexpression of CaHSP26 in transgenic tobacco alleviates photoinhibition of PSII and PSI during chilling stress under low irradiance. J. Plant Physiol. 2007, 164, 126–136. [Google Scholar] [CrossRef]
- Wang, M.L.; Zou, Z.W.; Li, Q.H.; Xin, H.H.; Zhu, X.J.; Chen, X.; Li, X.H. Heterologous expression of three Camellia sinensis small heat shock protein genes confers temperature stress tolerance in yeast and Arabidopsis thaliana. Plant Cell Rep. 2017, 36, 1125–1135. [Google Scholar] [CrossRef] [PubMed]
- Orvar, B.L.; Sangwan, V.; Omann, F.; Dhindsa, R.S. Early steps in cold sensing by plant cells: The role of actin cytoskeleton and membrane fluidity. Plant J. 2000, 23, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Knight, H.; Knight, M.R. Imaging spatial and cellular characteristics of low temperature calcium signature after cold acclimation in Arabidopsis. J. Exp. Bot. 2000, 51, 1679–1686. [Google Scholar] [CrossRef] [Green Version]
- Shukla, V.; Upadhyay, R.K.; Tucker, M.L.; Giovannoni, J.J.; Rudrabhatla, S.V.; Mattoo, A.K. Transient regulation of three clustered tomato class-I small heat-shock chaperone genes by ethylene is mediated by SlMADS-RIN transcription factor. Sci. Rep. 2017, 7, 6474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J. Analysis of SlHSP20 Gene Cluster Characteristics and SlHSP17.6A Gene Function under Cold Stress in Tomato. Master’s Thesis, Shenyang Agricultural University, Shenyang, China, June 2020. [Google Scholar]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.L.; Shi, Y.T.; Yang, S.H. Molecular mechanisms of plant responses to cold stress. Chin. Bull. Life Sci. 2015, 27, 398–405. [Google Scholar]
- Li, X.Y.; Wang, J.Q.; Bu, D.P.; Wei, H.Y.; Hu, H.; Zhou, L.Y. Advanced Research of Effect of Polyunsaturated Fatty Acidson Cell Membrane Function. Biotechnol. Bull. 2009, 12, 22–26. [Google Scholar] [CrossRef]
- Velly, H.; Bouix, M.; Passot, S.; Penicaud, C.; Beinsteiner, H.; Ghorbal, S.; Lieben, P.; Fonseca, F. Cyclopropanation of unsaturated fatty acids and membrane rigidification improve the freeze- drying resistance of Lact. Lact. Subsp. Lact. TOMSC161. Appl. Microbiol. Biotechnol. 2015, 99, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Miquel, M.; James, D.; Dooner, H., Jr.; Browse, J. Arabidopsis requires polyunsaturated lipids for low-temperature survival. Proc. Natl. Acad. Sci. USA 1993, 90, 6208–6212. [Google Scholar] [CrossRef] [Green Version]
- Tsvetkova, N.M.; Horváth, I.; Török, Z.; Wolkers, W.F.; Balogi, Z.; Shigapova, N.; Crowe, L.M.; Tablin, F.; Vierling, E.; Crowe, J.H. Small heat-shock proteins regulate membrane lipid polymorphism. Proc. Natl. Acad. Sci. USA 2002, 99, 13504–13509. [Google Scholar] [CrossRef] [Green Version]
- Balogi, Z.; Torok, Z.; Balogh, G.; Josvay, K.; Shigapova, N.; Vierling, E.; Vígh, L.; Horváth, I. “Heat shock lipid” in cyanobacteria during heat/light-acclimation. Arch. Biochem. Biophys. 2005, 436, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Maanni, A.; Dubertret, G.; Delrieu, M.J.; Roche, O.; Trémolières, A. Mutants of Chlamydomonas reinhardtii affected in phosphatidylglycerol metabolism and thylakoid biogenesis. Plant Physiol. Biochem. 1998, 36, 609–619. [Google Scholar] [CrossRef]
- Dubertret, G.; Gerard-Hirne, C.; Trémolières, A. Importance of trans-Δ3-hexadecenoic acid-containing phosphatidylglycerol in the formation of the trimeric light-harvesting complex in Chlamydomonas. Plant Physiol. Biochem. 2002, 40, 829–836. [Google Scholar] [CrossRef]
- Kruse, O.; Hankamer, B.; Konczak, C.; Gerle, C.; Morris, E.; Radunz, A.; Schmid, G.H.; Barber, J. Phosphatidylglycerol is involved in the dimerization of photosystem II. J. Biol. Chem. 2000, 275, 6509–6514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagio, M.; Gombos, Z.; Varkonyi, Z.; Masmaoto, K.; Sato, N.; Tsuzuki, M.; Wada, H. Direct evidence for requirement of PhosPhatidylglycerol in photosystem II of photosynthesis. Plant Physiol. 2000, 124, 795–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, N.; Hagio, M.; Wada, H.; Tsuzuki, M. Requirement of phosphatidylglycerol for Photosynthetic function in thylakoid membranes. Proc. Natl. Acad. Sci. USA 2000, 97, 10655–10660. [Google Scholar] [CrossRef] [Green Version]
- Gombos, Z.; Várkonyi, Z.; Hagio, M.; Iwaki, M.; Kovács, L.; Masamoto, K.; Itoh, S.; Wada, H. PhosPhatidylglycerol requirement for the function of electron acceptor plantoquinone QB in the photosystym II reaction center. Biochemistry 2002, 41, 3796–3802. [Google Scholar] [CrossRef]
- Xu, C.C.; Hartel, H.; Wada, H.; Hagio, M.; Yu, B.; Eakin, C.; Benning, C. The pgp1 Mutant locus of Arabidopsis encodes a phosphatidylglycerolphosphate synthase with impaired activity. Plant Physiol. 2002, 129, 594–604. [Google Scholar] [CrossRef] [Green Version]
- Hagio, M.; Sakurai, I.; Sato, S.; Kato, T.; Tabata, S.; Wada, H. Phosphatidylglycerol is essential for the development of thylakoid membranes in Arab. Thalina. Plant Cell Physiol. 2002, 43, 1456–1464. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.H. Effects of Low Temperatue and Weak Light on Photoinhibition and Thylakoid Membrane Lipids in Cucumber Leaves. Master’s Thesis, Shandong Agricultral University, Shandong, China, May 2003. [Google Scholar]
- Garnier, J.; Wu, B.; Maroc, J.; Guyon, D.; Trémolières, A. Restoration of both an oligoimeric form of the light-harvesting antenna CPII and a fluorescence state II-state I transition by Δ3-trans-hexadecenoic acid containing phosphatidylglycerol in cells of a mutant of chlamydomonas reinhardtii. Biochim. Biophys. Acta—Bioenerg. 1990, 1020, 153–162. [Google Scholar] [CrossRef]
- Wada, H.; Murata, N. The essential role of phosphatidylglycerol in photosynthesis. Photosynth. Res. 2007, 92, 205–215. [Google Scholar] [CrossRef]
- Li, Q.; Shen, W.Y.; Mavraganis, L.; Wang, L.P.; Gao, P.; Gao, J.; Cram, D.; Li, Y.F.; Liu, Z.Y.; Fowler, D.B.; et al. Elucidating the biochemical basis of trans-16:1 fatty acid change in leaves during cold acclimation in wheat. Plant Environ. Interact. 2021, 2, 101–111. [Google Scholar] [CrossRef]
- Ding, Y.L.; Shi, Y.T.; Yang, S.H. Molecular regulation of plant responses to environmental temperatures. Mol. Plant 2020, 13, 544–564. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.L.; Yang, S.H. Surviving and thriving: How plants perceive and respond to temperature stress. Dev. Cell. 2022, 57, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Carpaneto, A.; Ivashikina, N.; Levchenko, V.; Krol, E.; Jeworutzki, E.; Zhu, J.K.; Hedrich, R. Cold transiently activates calcium-permeable channels in Arabidopsis mesophyll cells. Plant Physiol. 2007, 143, 487–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penfield, S. Temperature perception and signal transduction in plants. New Phyiol. 2008, 179, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Sangwan, V.; Foulds, I.; Singh, J.; Dhindsa, R.S. Cold activation of Brassica napus BN115 promoter is mediated by structural changes in membranes and cytoskeleton, and requires Ca2+ in flux. Plant J. 2001, 27, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.L.; Shi, Y.T.; Yang, S.H. Advances and challenges in uncovering cold tolerance regulatory mechanisms in plants. New Phytol. 2019, 222, 1690–1704. [Google Scholar] [CrossRef] [Green Version]
- Chinnusamy, V.; Zhu, J.; Zhu, J.K. Cold stress regulation of gene expression in plants. Trends Plant Sci. 2007, 12, 444–451. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Zhu, J.K.; Sunkar, R. Gene regulation during cold stress acclimation in plants. Methods Mol. Biol. 2010, 639, 39–55. [Google Scholar] [CrossRef] [Green Version]
- Fowler, S.; Thomashow, M.F. Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway. Plant Cell. 2002, 14, 1675–1690. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, T.H.; Lee, J.T.; Yang, P.T.; Chiu, L.H.; Charng, Y.Y.; Wang, Y.C.; Chan, M.T. Heterology expression of the Arabidopsis C-repeat/dehydration response element binding factor 1 gene confers elevated tolerance to chilling and oxidative stresses in transgenic tomato. Plant Physiol. 2002, 129, 1086–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Rathore, M.; Goyary, D.; Singh, R.K.; Anandhan, S.; Sharma, D.K.; Ahmed, Z. Induced ectopic expression of At-CBF1 in marker-free transgenic tomatoes confers enhanced chilling tolerance. Plant Cell Rep. 2011, 30, 1019–1028. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Yang, J.S.; Guo, S.J.; Meng, J.J.; Zhang, Y.L.; Wan, S.B.; He, Q.W.; Li, X.G. Over-expression of the Arabidopsis CBF1 gene improves resistance of tomato leaves to low temperature under low irradiance. Plant Biol. 2011, 13, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.D.; Shi, Y.; Zhu, N.; Zhong, S.L.; Bouzayen, M.; Li, Z.G. SlGRAS4 mediates a novel regulatory pathway promoting chilling tolerance in tomato. Plant Biotechnol. J. 2020, 18, 1620–1633. [Google Scholar] [CrossRef] [PubMed]
- Mu, C.; Zhang, S.; Yu, G.; Chen, N.; Li, X.; Liu, H. Overexpression of small heat shock protein LimHSP16.45 in Arabidopsis enhances tolerance to abiotic stresses. PLoS ONE 2013, 8, e82264. [Google Scholar] [CrossRef] [Green Version]
- Dean, M.E.; Johnson, J.L. Human Hsp90 cochaperones: Perspectives on tissue-specific expression and identification of cochaperones with similar in vivo functions. Cell Stress Chaperones 2021, 1, 3–13. [Google Scholar] [CrossRef]
- Sato, S.; Chieri, T.S.; Amol, R.; Ronald, C.C.; Joan, W.C. Structure and functions of the Mediator complex: A key regulator of RNA polymerase II transcription. FASEB J. 2020, 34, 121. [Google Scholar] [CrossRef]
- Borggrefe, T.; Yue, X. Interactions between subunits of the Mediator complex with gene-specific transcription factors. Semin. Cell Dev. Biol. 2011, 22, 759–768. [Google Scholar] [CrossRef]
- Allen, B.L.; Taatjes, D.J. The Mediator complex: A central integrator of transcription. Nat. Rev. Mol. Cell Biol. 2015, 16, 155–166. [Google Scholar] [CrossRef]
- Piers, A.H.; Charlotte, H.H.; Ewon, K.; Rebecca, L.; Marc, R.K.; Elizabeth, A.D.C.; John, F.S.; Heather, K. The Arabidopsis mediator complex subunits MED16, MED14, and MED2 regulate mediator and RNA polymerase II recruitment to CBF-responsive cold-regulated genes. Plant Cell 2014, 26, 465–484. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.H.; Duan, X.F.; Wang, S.; Wu, W.Y.; Zhang, X.C. Virus-induced gene silencing (VIGS) for functional analysis of MYB80 gene involved in Solanum lycopersicum cold tolerance. Protoplasma 2019, 256, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.P.; Pan, J.W.; Zhang, M.Y.; Xing, X.; Zhou, Y.; Liu, Y.; Li, D.P.; Li, D.Q. ZmMKK4, a novel group C mitogen-activated protein kinase kinase in maize (Zea mays), confers salt and cold tolerance in transgenic Arabidopsis. Plant Cell Environ. 2011, 34, 1291–1303. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhang, J. Effect of abscisic acid on active oxygen species, antioxidative defence system and oxidative damage in leaves of maize seedlings. Plant Cell Physiol. 2001, 42, 1265–1273. [Google Scholar] [CrossRef]
- Zhang, A.; Jiang, M.; Zhang, J.; Tan, M.; Hu, X. Mitogen-activated protein kinase is involved in abscisic acid-induced antioxidant defense and acts downstream of reactive oxygen species production in leaves of maize plants. Plant Physiol. 2006, 141, 475–487. [Google Scholar] [CrossRef] [Green Version]
- Rao, M.V.; Paliyath, C.; Ormrod, D.P. Ultraviolet-B- and ozone induced biochemical changes in antioxidant enzymes of Arabidopsis thaliana. Plant Physiol. 1996, 110, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Kong, F.Y.; Deng, Y.S.; Zhou, B.; Wang, G.D.; Wang, Y.; Meng, Q.W. A chloroplast-targeted DnaJ protein contributes to maintenance of photosystem II under chilling stress. J. Exp. Bot. 2014, 65, 143–158. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.H.; Jaafar, H.Z.E. Primary, secondary metabolites, H2O2, malondialdehyde and photosynthetic responses of Orthosiphon stimaneus Benth. to different irradiance levels. Molecules 2012, 17, 1159–1176. [Google Scholar] [CrossRef]
- Quan, R.; Shang, M.; Zhang, H.; Zhao, Y.; Zhang, J. Improved chilling tolerance by transformation with betA gene for the enhancement of glycinebetaine synthesis in maize. Plant Sci. 2004, 166, 141–149. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, D.C.; Miao, Q.S.; Yang, J.; Xuan, Y.H.; Hu, Y.B. Essential role of sugar transporter OsSWEET11 during the early stage of rice grain filling. Plant Cell Physiol. 2017, 58, 863–873. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.S.; Feng, C.Y.; Wang, M.N.; Li, T.L.; Liu, X.; Jiang, J. Plasma membrane-localized SlSWEET7a and SlSWEET14 regulate sugar transport and storage in tomato fruits. Hortic. Res. 2021, 8, 186–202. [Google Scholar] [CrossRef]
- Zhao, T.; Yang, H.; Jiang, J.; Liu, G.; Zhang, H.; Xiao, D.; Chen, X.; Li, J.; Xu, X. Silencing of the SAMDC gene decreases resistance of tomato to Cladosporium fulvum. Physiol. Mol. Plant Pathol. 2018, 102, 1–7. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | ID | MS2 Metabolite | Log2FC (0 h) | Log2FC (12 h) |
|---|---|---|---|---|
| Glycerophospholipids | M719T496 | PG 32:1; PG (16:0/16:1) | +7.08 | +4.31 |
| M518T391 | LysoPS 18:3; LysoPS 18:3 | +3.34 | ||
| M571T536_2 | LysoPI 16:0; LysoPI 16:0 | −0.44 | ||
| M695T527 | PA 36:4; PA (18:2/18:2) | −0.32 | ||
| M781T462 | 1-Palmitoyl-2-linoleoyl-sn-glycero-3-phosphocholine | −0.04 | ||
| M781T493 | 1-Palmitoyl-2-linoleoyl-sn-glycero-3-phosphocholine | −0.08 | ||
| M781T533 | 1-Palmitoyl-2-linoleoyl-sn-glycero-3-phosphocholine | −0.21 | ||
| M184T59 | Phosphocholine | −0.08 | ||
| Glycerolipids | M815T497 | SQDG 34:3; SQDG (16:0/18:3) | −0.45 | |
| M830T521 | SQDG 35:3; SQDG (17:0/18:3) | −0.23 | −0.33 | |
| Fatty acids | M279T422 | 2-2-Hexyldecanoic acid | +2.13 | |
| M293T414 | 9-Oxo-10E,12Z,15Z-octadecatrienoic acid | +2.02 | ||
| M328T288 | 9-Oxo-11-(3-pentyl-2-oxiranyl)-10E-undecenoic acid | +2.41 | ||
| M328T346 | 9-Oxo-11-(3-pentyl-2-oxiranyl)-10E-undecenoic acid | +2.44 | ||
| M265T377 | Dinor-12-oxophytodienoic acid | +2.12 | ||
| M353T393 | Monolinolenin (9c,12c,15c) | +2.26 | ||
| M145T53 | (E)-2-Methylglutaconic acid | −0.36 | ||
| M337T316 | 8,11-Tridecadienoic acid, 13-(3-pentyl-2-oxiranyl)-, methyl ester, (8Z,11Z)- | −0.23 | ||
| M221T231 | 2,3-Dimethyl-3-hydroxyglutaric acid | +3.08 | ||
| M293T400_2 | 9-OxoODE | +2.00 | ||
| M311T353 | (E)-13-Hydroxy-10-oxo-11-octadecenoic acid | +2.53 | ||
| M275T400 | Stearidonic acid | +2.35 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Lv, S.; Zhao, Y.; Chang, C.; Hong, W.; Jiang, J. SlHSP17.7 Ameliorates Chilling Stress-Induced Damage by Regulating Phosphatidylglycerol Metabolism and Calcium Signal in Tomato Plants. Plants 2022, 11, 1865. https://doi.org/10.3390/plants11141865
Wu Y, Lv S, Zhao Y, Chang C, Hong W, Jiang J. SlHSP17.7 Ameliorates Chilling Stress-Induced Damage by Regulating Phosphatidylglycerol Metabolism and Calcium Signal in Tomato Plants. Plants. 2022; 11(14):1865. https://doi.org/10.3390/plants11141865
Chicago/Turabian StyleWu, Yuanyuan, Shuwen Lv, Yaran Zhao, Chenliang Chang, Wei Hong, and Jing Jiang. 2022. "SlHSP17.7 Ameliorates Chilling Stress-Induced Damage by Regulating Phosphatidylglycerol Metabolism and Calcium Signal in Tomato Plants" Plants 11, no. 14: 1865. https://doi.org/10.3390/plants11141865