
Immunohistochemical Distribution of Serotonin Transporter (SERT) in the Optic Lobe of the Honeybee, Apis mellifera

 ,
,  , , , and
, , , and 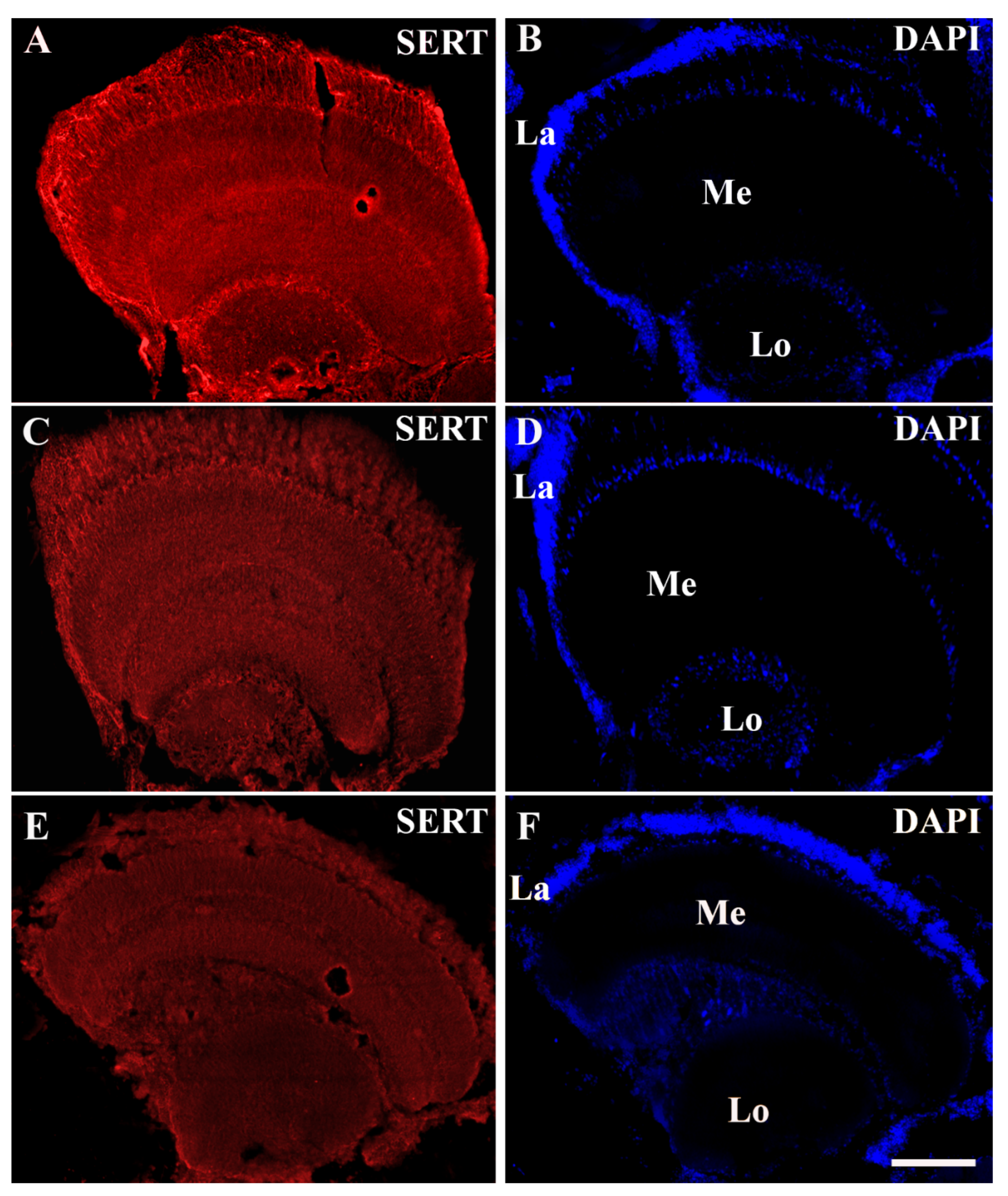{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Fixation
2.2. Immunostaining
2.3. Analysis of Sections
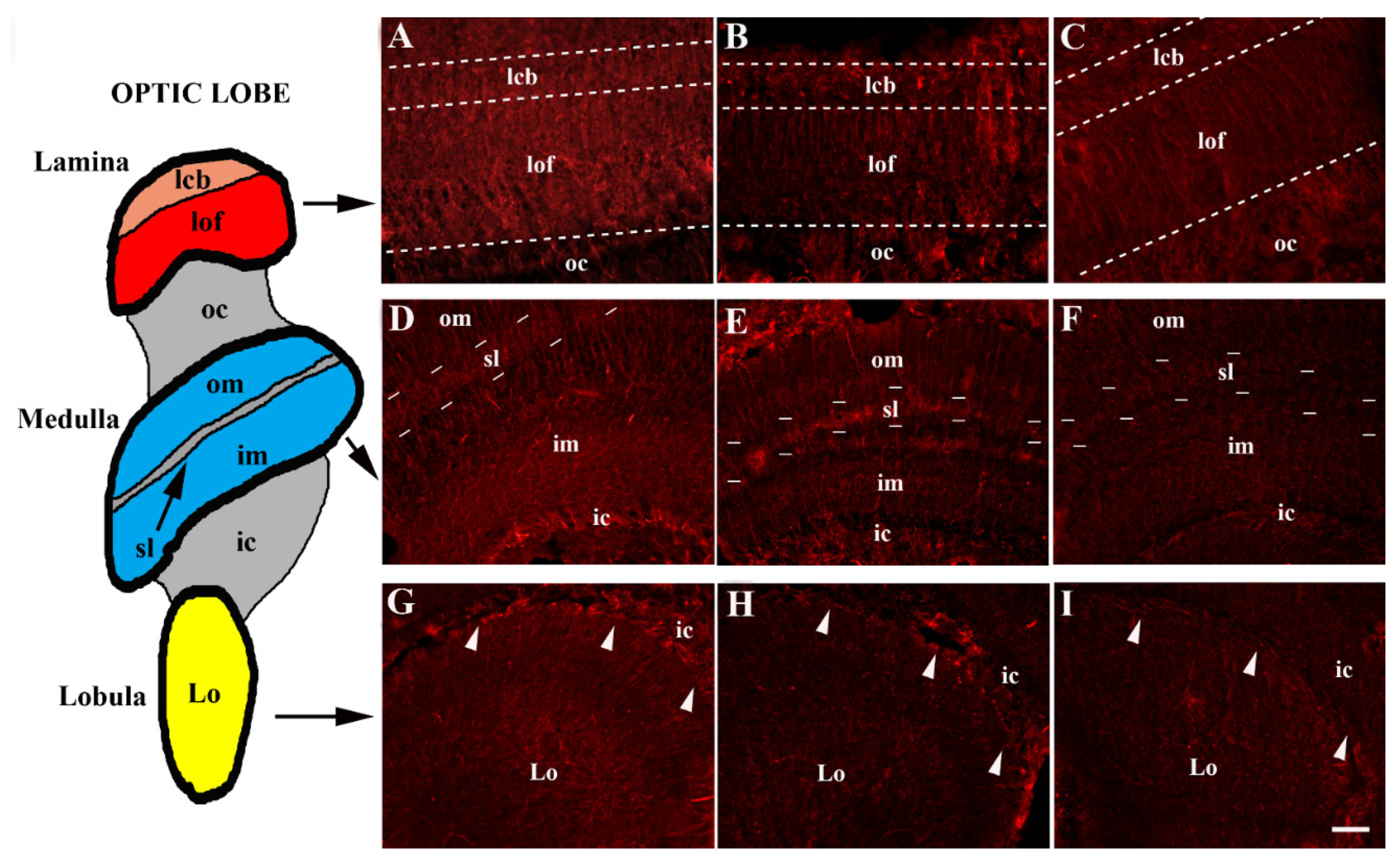
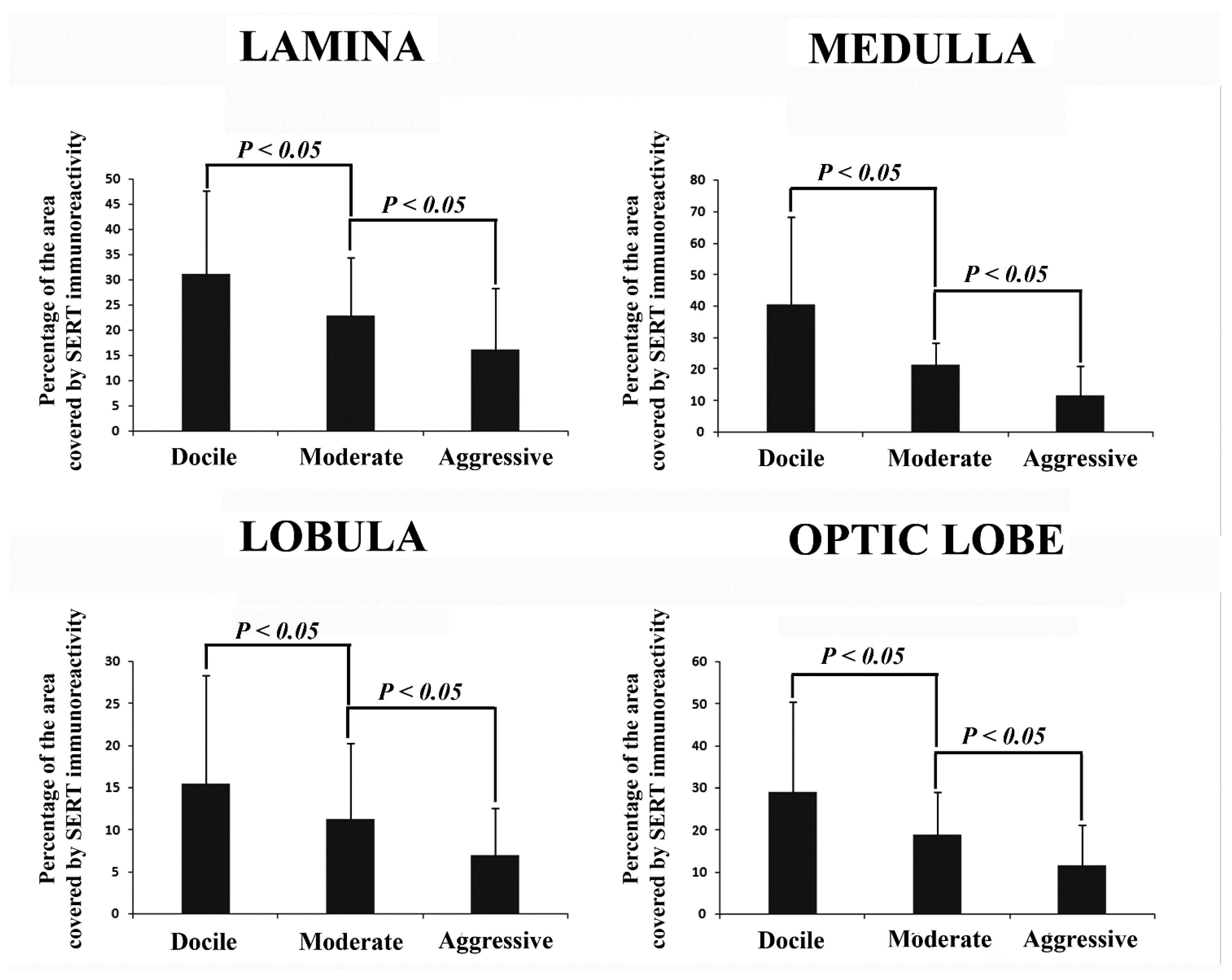
3. Results
Distribution of SERT Immunoreactivity in the Optic Lobe
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ito, K.; Shinomiya, K.; Ito, M.; Armstrong, J.D.; Boyan, G.; Hartenstein, V.; Harzsch, S.; Heisenberg, M.; Homberg, U.; Jenett, A.; et al. A systematic nomenclature for the insect brain. Neuron 2014, 81, 755–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dierick, H.A.; Greenspan, R.J. Serotonin and neuropeptide F have opposite modulatory effects on fly aggression. Nat. Genet. 2007, 39, 678–682. [Google Scholar] [CrossRef] [PubMed]
- Sitaraman, D.; Zars, M.; LaFerriere, H.; Chen, Y.-C.; Sable-Smith, A.; Kitamoto, T.; Rottinghaus, G.E.; Zars, T. Serotonin is necessary for place memory in drosophila. Proc. Natl. Acad. Sci. USA 2008, 105, 5579–5584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheiner, R.; Baumann, A.; Blenau, W. Aminergic control and modulation of honeybee behaviour. Curr. Neuropharmacol. 2006, 4, 259–276. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.A.; Krantz, D.E. Drosophila melanogaster as a genetic model system to study neurotransmitter transporters. Neurochem. Int. 2014, 73, 71–88. [Google Scholar] [CrossRef] [Green Version]
- Tierney, A.J. Invertebrate serotonin receptors: A molecular perspective on classification and pharmacology. J. Exp. Biol. 2018, 221, jeb184838. [Google Scholar] [CrossRef] [Green Version]
- Moutkine, I.; Collins, E.L.; Béchade, C.; Maroteaux, L. Evolutionary considerations on 5-HT2 receptors. Pharmacol. Res. 2019, 140, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Brandt, R.; Rohlfing, T.; Rybak, J.; Krofczik, S.; Maye, A.; Westerhoff, M.; Hege, H.-C.; Menzel, R. Three-dimensional average-shape atlas of the honeybee brain and its applications. J. Comp. Neurol. 2005, 492, 1–19. [Google Scholar] [CrossRef]
- Paulk, A.C.; Stacey, J.A.; Pearson, T.W.J.; Taylor, G.J.; Moore, R.J.D.; Srinivasan, M.V.; van Swinderen, B. Selective attention in the honeybee optic lobes precedes behavioral choices. Proc. Natl. Acad. Sci. USA 2014, 111, 5006–5011. [Google Scholar] [CrossRef] [Green Version]
- Schürmann, F.W.; Klemm, N. Serotonin-immunoreactive neurons in the brain of the honeybee. J. Comp. Neurol. 1984, 225, 570–580. [Google Scholar] [CrossRef]
- Thamm, M.; Balfanz, S.; Scheiner, R.; Baumann, A.; Blenau, W. Characterization of the 5-HT1A receptor of the honeybee (Apis mellifera) and involvement of serotonin in phototactic behavior. Cell. Mol. Life Sci. 2010, 67, 2467–2479. [Google Scholar] [CrossRef] [PubMed]
- Thamm, M.; Rolke, D.; Jordan, N.; Balfanz, S.; Schiffer, C.; Baumann, A.; Blenau, W. Function and distribution of 5-HT2 receptors in the honeybee (Apis mellifera). PLoS ONE 2013, 8, e82407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erber, J.; Kloppenburg, P. The modulatory effects of serotonin and octopamine in the visual system of the honey bee (Apis mellifera L.). J. Comp. Physiol. A 1995, 176, 119–129. [Google Scholar] [CrossRef]
- Seidel, C.; Bicker, G. The developmental expression of serotonin-immunoreactivity in the brain of the pupal honeybee. Tissue Cell 1996, 28, 663–672. [Google Scholar] [CrossRef]
- Nielsen, K.; Brask, D.; Knudsen, G.M.; Aznar, S. Immunodetection of the serotonin transporter protein is a more valid marker for serotonergic fibers than serotonin. Synapse 2006, 59, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Buchler, R.; Andonov, S.; Bienefeld, K.; Costa, C.; Hatjina, F.; Kezic, N.; Kryger, P.; Pivak, M.; Uzunov, A.; Wilde, J. Standard methods for rearing and selection of Apis mellifera queens. J. Apic. Res. 2013, 52, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Uzunov, A.; Büchler, R.; Bienefeld, K. Performance Testing Protocol. A Guide for European Honey Bee Breeders, 1st ed.; Landesbetrieb Landwirtschaft Hessen: Kirchhain, Germany, 2015. [Google Scholar]
- Uzunov, A.; Brascamp, E.W.; Buchler, R. The basic concept of honey bee breeding programs. Bee World 2017, 94, 84–87. [Google Scholar] [CrossRef]
- Hoppe, A.; Du, M.; Bernstein, R.; Tiesler, F.K.; Kärcher, M.; Bienefeld, K. Substantial genetic progress in the international Apis mellifera carnica population since the implementation of genetic evaluation. Insects 2020, 11, 768. [Google Scholar] [CrossRef]
- Kravitz, E.A.; Huber, R. Aggression in invertebrates. Curr. Opin. Neurobiol. 2003, 13, 736–743. [Google Scholar] [CrossRef]
- Bubak, A.N.; Watt, M.J.; Yaeger, J.D.W.; Renner, K.J.; Swallow, J.G. The stalk-eyed fly as a model for aggression—Is there a conserved role for 5-HT between vertebrates and invertebrates? J. Exp. Biol. 2020, 223, jeb132159. [Google Scholar] [CrossRef]
- Poetini, M.R.; Musachio, E.A.S.; Araujo, S.M.; Almeida, F.P.; Dahleh, M.M.M.; Bortolotto, V.C.; Janner, D.E.; Pinheiro, F.C.; Ramborger, B.P.; Roehrs, R.; et al. Iron overload during the embryonic period develops hyperactive like behavior and dysregulation of biogenic amines in drosophila melanogaster. Dev. Biol. 2021, 475, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Tedjakumala, S.R.; Aimable, M.; Giurfa, M. Pharmacological modulation of aversive responsiveness in honey bees. Front. Behav. Neurosci. 2014, 7, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birmingham, J.T.; Tauck, D.L. Neuromodulation in invertebrate sensory systems: From biophysics to behavior. J. Exp. Biol. 2003, 206, 3541–3546. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bombardi, C.; Salamanca, G.; Tagliavia, C.; Grandis, A.; Mille, F.; De Iorio, M.G.; Minozzi, G. Immunohistochemical Distribution of Serotonin Transporter (SERT) in the Optic Lobe of the Honeybee, Apis mellifera. Animals 2022, 12, 2032. https://doi.org/10.3390/ani12162032
Bombardi C, Salamanca G, Tagliavia C, Grandis A, Mille F, De Iorio MG, Minozzi G. Immunohistochemical Distribution of Serotonin Transporter (SERT) in the Optic Lobe of the Honeybee, Apis mellifera. Animals. 2022; 12(16):2032. https://doi.org/10.3390/ani12162032
Chicago/Turabian StyleBombardi, Cristiano, Giulia Salamanca, Claudio Tagliavia, Annamaria Grandis, Fanny Mille, Maria Grazia De Iorio, and Giulietta Minozzi. 2022. "Immunohistochemical Distribution of Serotonin Transporter (SERT) in the Optic Lobe of the Honeybee, Apis mellifera" Animals 12, no. 16: 2032. https://doi.org/10.3390/ani12162032