
A Review on Environmental Contaminants-Related Fertility Threat in Male Fishes: Effects and Possible Mechanisms of Action Learned from Wildlife and Laboratory Studies

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. An Introduction to Reproductive Biology in Male Fishes
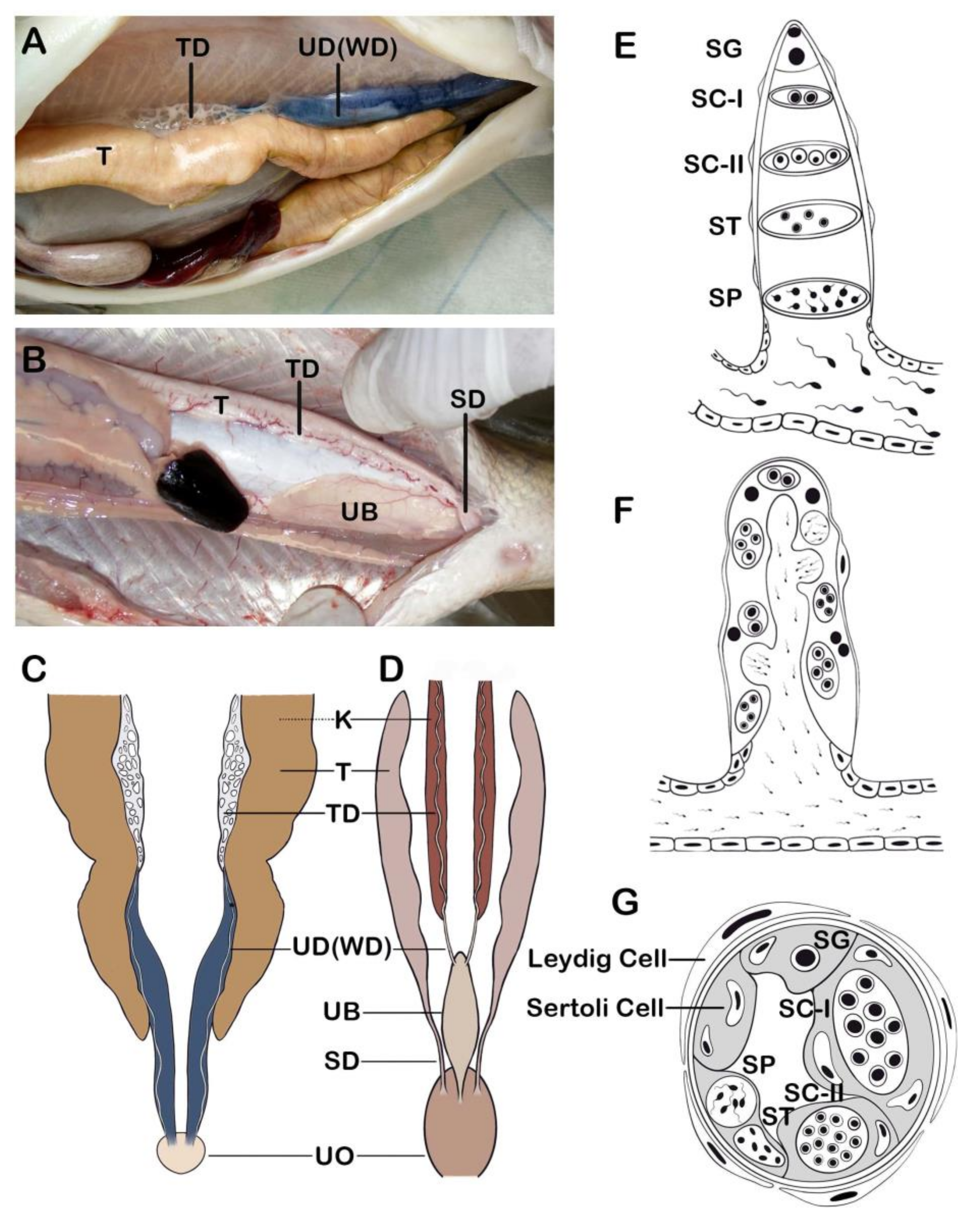
2.1. Anatomy of Reproductive Organ
2.2. Spermatogenesis
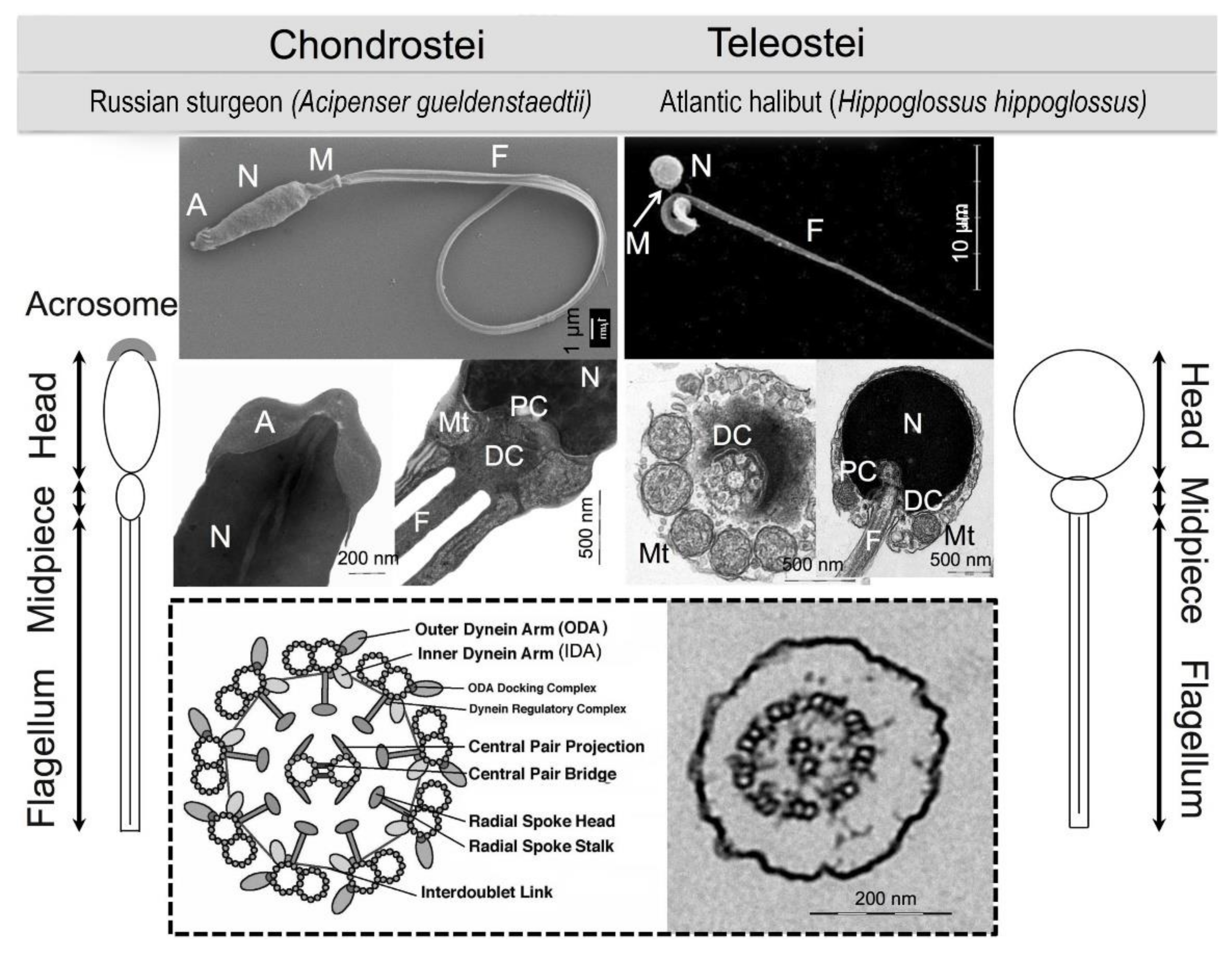
2.3. Sperm Morphology
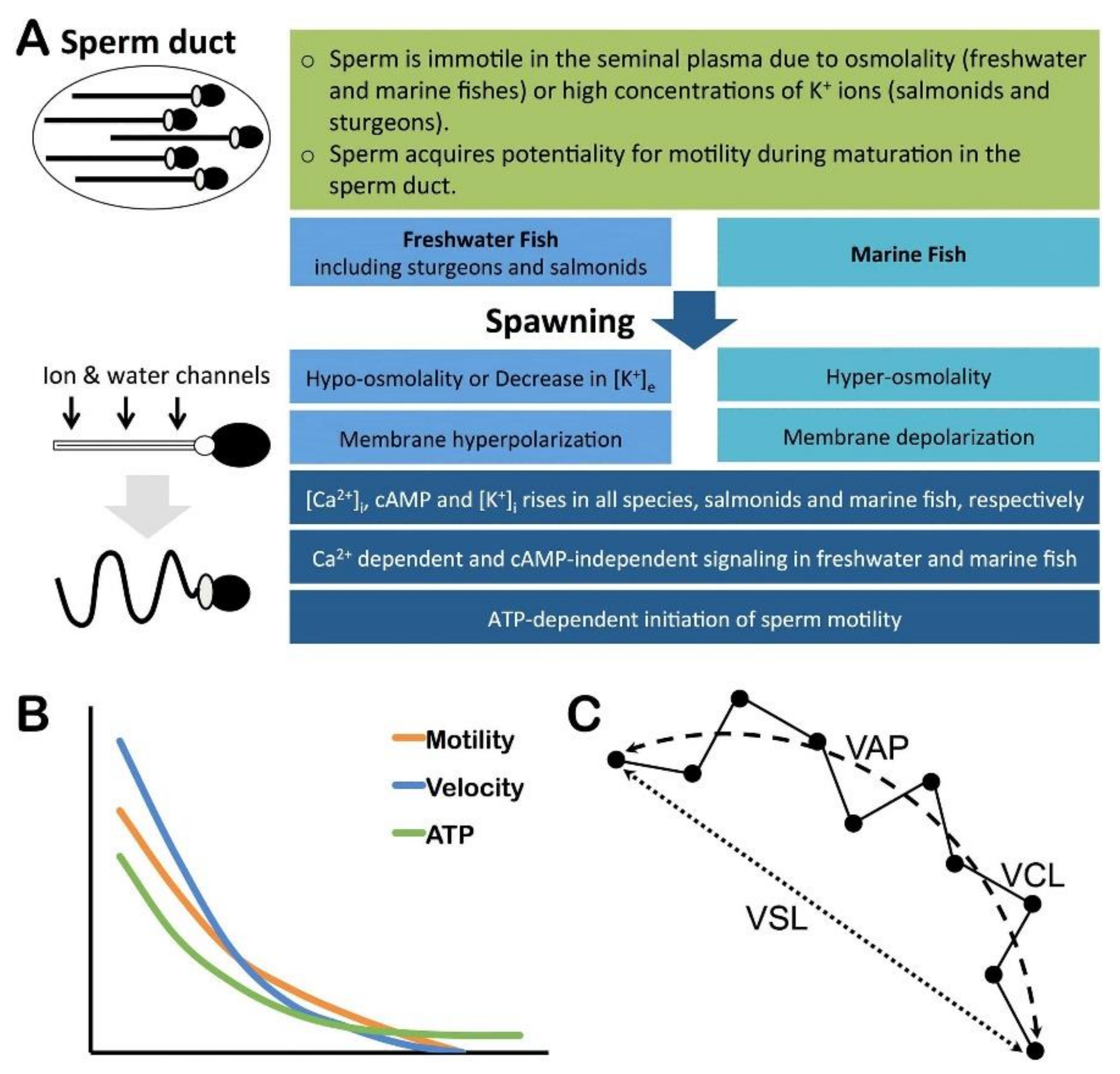
2.4. Sperm Physiology
2.5. Sperm Motility
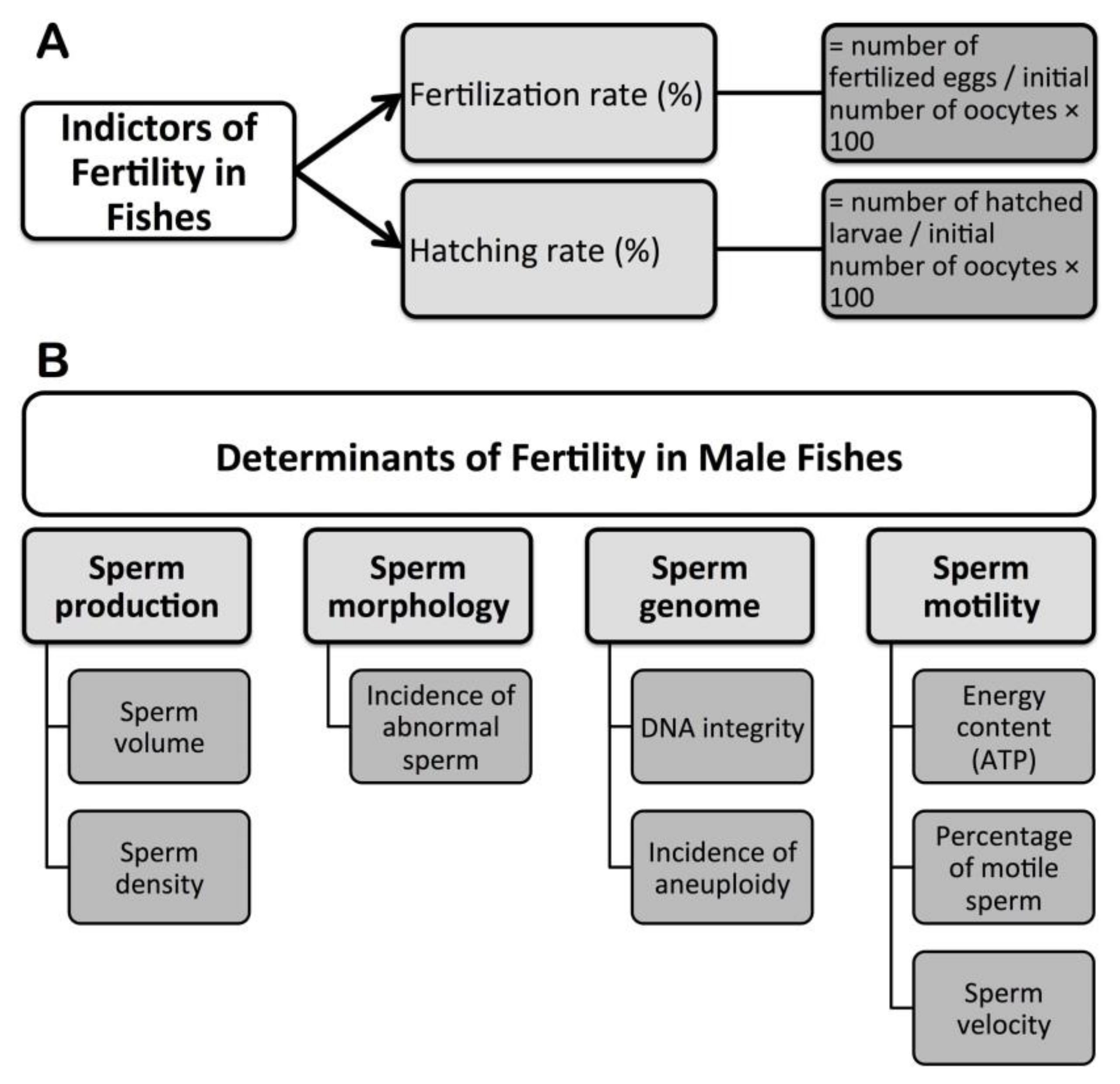
3. Fertility Indicators and Assessments in Fishes
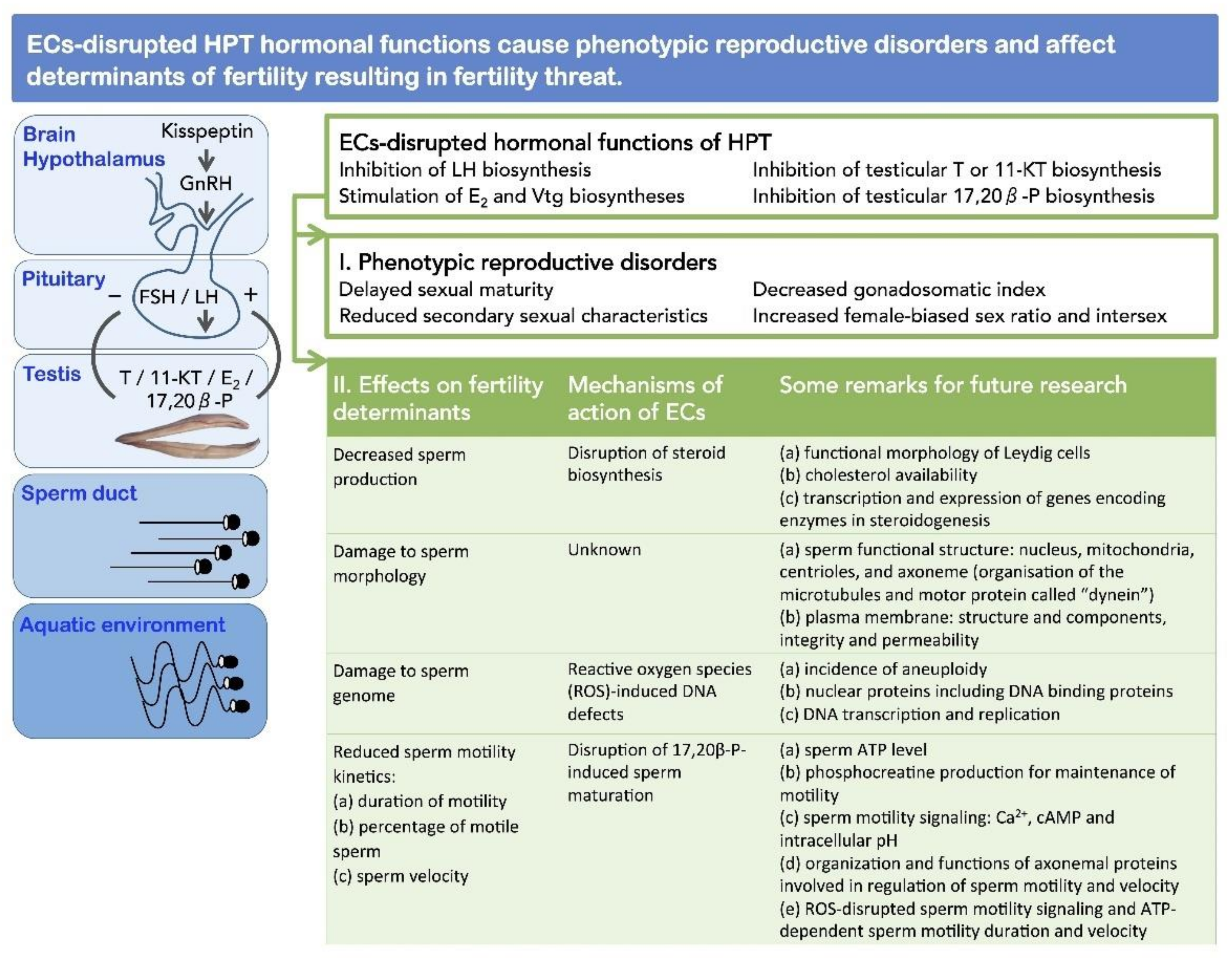
4. Determinants of Fertility in Male Fishes
4.1. Sperm Production
4.2. Sperm Morphology
4.3. Sperm Genome
4.4. Sperm Motility Kinetics
5. Wildlife Evidences for Environmental Contaminants (ECs)-Related Fertility Threat in Male Fishes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
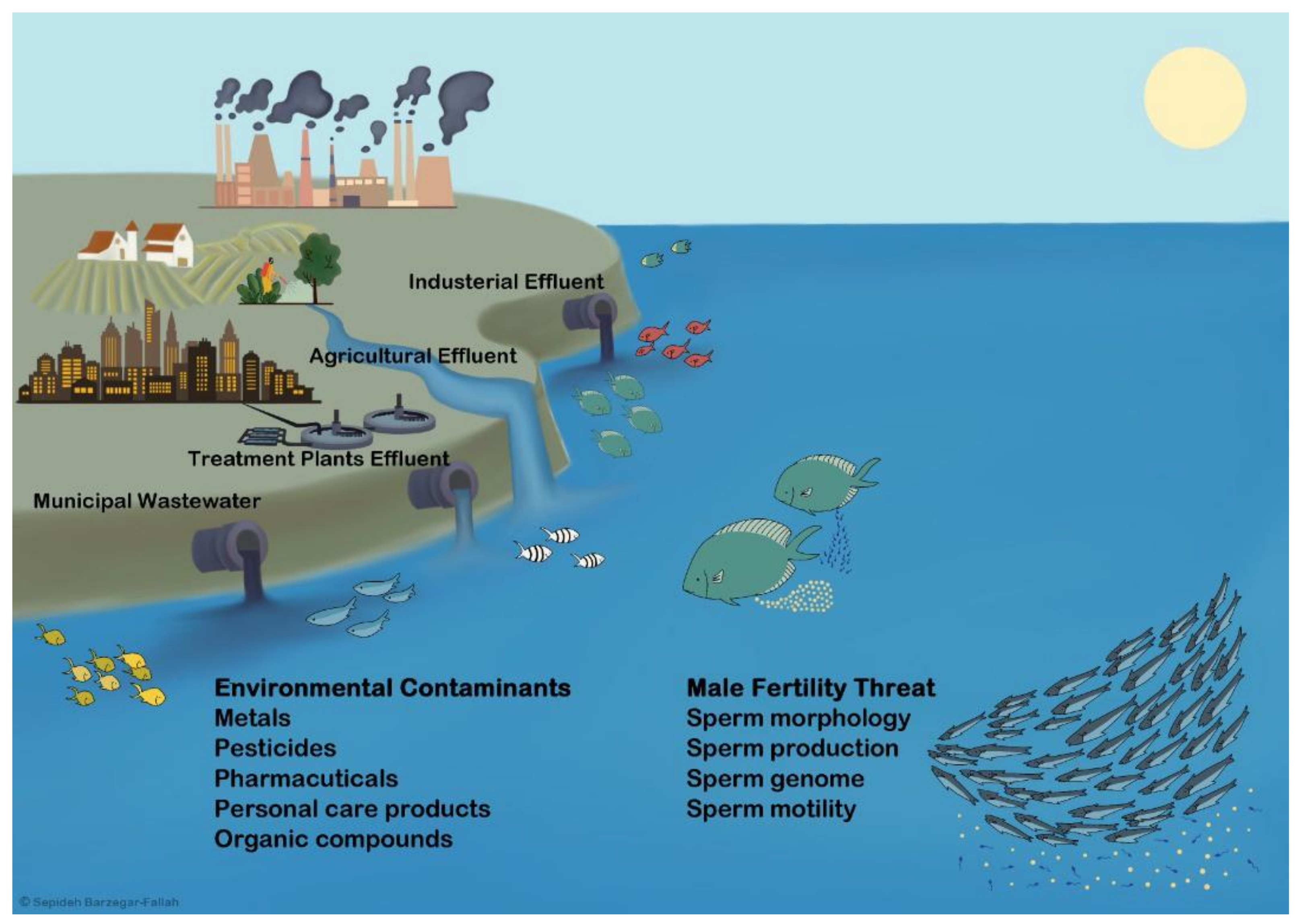{kind=link}
{kind=link}
| Region | Fish Species | Contaminants | Reproductive Endpoints | Description | Authors |
|---|---|---|---|---|---|
| Jackfish Bay, North Shore of Lake Superior, Canada | White sucker | BKME that includes resin acid | Delayed sexual maturation Decreased testicular size Reduced secondary sexual characters Decreased LH, T, 11-KT, and 17,20β-P levels Failed to increase 17,20β-P levels in response to stimulation of spermiation by sGnRH-A | Polluted site: Jackfish bay Reference site: Mountain bay Sampling time: May–Aug 1988 (Munkittrick et al. 1991); Aug 1988, Aug 1989 and Sep 1990 (Munkittrick et al. 1992); May 1990 and 1991 (Van Der Kraak et al. 1992) | [109,110,138] |
| Estuarine, UK | Flounder | Sewage effluents | No difference in GSI, increased Vtg levels at all polluted sites Intersex at the Mersey estuary (17%) | Polluted sites: Tyne, Crouch, Thames and Mersey estuaries Reference site: Alde River Sampling time: Sep–Dec 1996 | [111] |
| The Moselle and the Rhone Watershed, France | Chub | Metals and organic chemicals | No difference in GSI Increased Vtg levels at Igney Necrotic sperm cells observed at Igney | Polluted site: Igney (the Moselle river) Reference site: Saillans (the Drome river) Sampling time: June | [139] |
| Saint Louis, Missouri, USA | Shovelnose sturgeon | Chlordane, p,p’. DDE, PCBs | Intersex (29%) | Polluted site: Downstream of St. Louis, Prairie de Rocher, IL Reference site: Upstream of St. Louis, Davenport, IA Sampling time: Autumn | [112] |
| Tokyo Bay, Japan | Flounder | Sewage effluents that include Alkylphenol polyethoxylates and NP | Decreased GSI in males with ovo-testis Increased Vtg levels Intersex (15% of individuals) | Polluted site: Tokyo Bay, Japan Reference site: Shiriuchi, Hokkaido, Japan Concentrations (μg/L): Alkylphenol polyethoxylates (37.5), Nonylphenol: (0.2–1.1) Sampling time: Jan 1997–May 1998 | [113] |
| The Po River, Italy | Barbel | Not determined | Intersex (50%) | Polluted site: Downstream of the Po River Reference site: Upstream of the Po River Sampling time: April 1999 | [114] |
| Nene and Aire Rivers, UK | Roach | STWs | Decreased GSI Intersex at Nene and Aire (100%) (1–4% intersex at reference site) Delayed spermatogenesis Increased Vtg, T, and E2 levels in intersex males compared to normal and to intersex males of reference sitesNo difference in 11-KT levels between intersex males of polluted sites and reference sites Decreased number of spermiating males Decreased sperm volume and density Decreased sperm motility and velocity Decreased fertilizing ability of sperm | Polluted sites: Nene (Northamptonshire) and Aire (Yorkshire) Reference cites: Royal Canal (Ireland), Grantham Canal (Leicestershire) and a spring-fed lake (Lake Wartnaby, Leicestershire) Sampling time: October 1995 and 1996; May 1998, 1999, and 2000 | [115,132] |
| The St. Johns River | Largemouth bass | DDT and its metabolites, PCBs, high and low molecular weight PAHs, cyclodiene and chlorinated pesticides | At pre-spawning season (Sep 1996): Increased E2 levels at polluted sites Decreased 11-KT levels at Palatka and Julington Creek At Spawning season (February 1997): Decreased GSI at Palatka, increased E2 levels at Palatka and Green Cove, decreased E2 levels at Julington CreekDecreased 11-KT levels at all polluted sites | Polluted sites: Downstream of the St. Johns River: Palatka (North of Welaka), Green Cove (North of Palatka) and Julington Creek (North of Green Cove). The Palatka and Green Cove sites are representative of urban, industrial, and agricultural development. The Palatka site is near to a paper mill plant. The Julington Creek site receives discharges of wastewater and runoff from recreational boating marinas Reference site: Upstream of the St. Johns River Sampling time: September 1996 and February 1997 | [116] |
| The Anoia River, Spain | Carp | STW that includes estrogenic compounds | Decreased testicular size Occurrence of testicular atrophy Intersex (19%) Decreased T levels, Increased Vtg levels | Polluted sites: Downstream of the Anoia River Reference site: None Sampling time: January and March 2000 | [126] |
| The St. Lawrence River, Montreal, Canada | Spottail Shiner | STW that includes xenoestrogens | Intersexes at Îles de la Paix, Île Dorval, Îlet Vert and Île Beauregard (2.6, 15, 31, and 27, respectively) Increased Vtg mRNA levels at Îlet Beauregard, Îlet Vert, and Île Saint-Ours compared to the Îles de Boucherville, Île Dorval, Îles de la Paix, and Ottawa river Delayed spermatogenesis at Îlet Vert and Île Beauregard Decreased sperm density, motility, and velocity at Îlet Vert compared to the Îles de la Paix | Polluted site: The downstream sites of the St. Lawrence River (Îlet Vert, Île Beauregard and Île St. Ours) Reference sites: The upstream sites of the St. Lawrence River (Îles de la Paix, Île Dorval, Îles de Boucherville) and the Ottawa River Sampling time: June 1999–2002 | [117] |
| Hamilton Harbour, Western Lake Ontario, Canada | White Perch | Treated domestic sewage | Intersex (22–83%) Increased Vtg levels | Polluted site: The Cootes Paradise region Reference site: Deal Lake and hatchery fish Sampling time: August 1998 and 2000, October 1999, July and September 2002 | [118] |
| Boulder Creek and the South Platte River, Colorado, USA | White sucker | WWTP | Intersex Increased percentage of female compared to males (83% vs. 45%) | Polluted site: Boulder Creek downstream of the Boulder WWTP Reference site: A stream reach beginning 2 km upstream of the Boulder Sampling time: March May, October and November 2002 | [120] |
| Danish Streams in Aarhus County, Jutland, Denmark | Roach | Domestic STWs | Intersex (6.7–6.5% at polluted sites compared to 4.5–5% at reference sites) The highest intersex 26.5% was observed in the stream Kristrup Landkanal. | Polluted sites: Aarhus Brook, Egaa, and Kristrup Landkanal streams receive sewage effluent discharges from STW Reference sites: Lake Almind and Lake Ravn receiving no or small amounts of STW, respectively Sampling time: June and September 1999 | [119] |
| Boulder Creek, a Rributary of the South Platte River, Colorado, USA | WWTP that includes E2, EE2, estrone, BPA, NP | Decreased GSI Intersex (18–22%) Increased Vtg levels Decreased sperm abundance | Polluted site: Downstream of the Boulder WWTP Reference site: Upstream of the Boulder WWTP The Boulder WWTP uses filter/activated sludge treatment process with nitrification/denitrification and chlorination/dechlorination. The mean annual concentrations of NH3-N, NO3-N, biological oxygen demand, and total suspended solids in the WWTP effluent were 7, 12, 15, and 6 mg/L, respectively. At downstream of the WWTP outfall, NH3-N and NO3-N were 1 and 4 mg/L, respectively Concentrations (ng/L): E2 (1.6–2.1), EE2 (0.7–<2.0), estrone (36), BPA (2.5-35), NP (39–340) at polluted site, and E2 (<0.2–2.9), EE2 (<0.8–<2.0), estrone (<5), BPA (3.1–27), NP (<33–120) at reference site. Sampling time: Autumn 2003 and Spring 2004 | [127] | |
| Oldman, Bow and Red Deer Rivers, Alberta, Canada | Longnose dace | Municipal wastewater and agricultural runoff | Increased female-biased sex ratio at Oldman river Increased GSI at downstream of Oldman and Bow rivers Increased Vtg levels at downstream Oldman, Bow, and Red Deer rivers | Polluted sites: Downstream of Oldman, Bow and Red deer rivers (Discharges of Lethbridge, Calgary and Red Deer, respectively) Reference sites: Upstream of the above rivers Concentrations (ng/L) of selected contaminants at Red Deer and Oldman rivers: α-estradiol (0.2), E2 (0.6), T (1.7), Equilin (1.8), Estrone (1.8), BPA (3.8), α-Zearalanol (5.2), Ergosterol (25.4), Stigmastanol (37.5), β-sitosterol (100.5), Stigmasterol (144.7), Cholesterol (189.2), Fucosterol (212.1) Sampling time: April 2005 (Jeffries et al. 2008), Oct and No 2005 (Jeffries et al. 2010) | [123,124] |
| The Luvuvhu River, Limpopo Province, South Africa | Tilapia | DDT and metabolites | Decreased GSI at XW Intersex at AD, ND, and XW (55%, 60%, and 59%, respectively) Increased sperm velocity at ND Trends toward decrease in sperm motility at ND and XW | Polluted sites: Nandoni Dam (ND) and Xikundu Weir (XW) Reference site: Albasini Dam (AD) (outside of the DDT-sprayed area) Sampling surveys were carried out during high and low flow seasons Concentrations (μg/L): DDT and metabolites: 0.1–1 Sampling time: March and October 2007, February 2008 | [121,122] |
| African catfish | A trend toward decrease in sperm motility at ND Trends toward decrease in sperm velocity at ND and XW Testicular abnormalities at ND and XW | ||||
| The Elbe River, Czech Republic | Chub | Hg, PCBs, DDT and its metabolites, HCHs, HCB, OCS | Intersex at downstream of Usti nad Labem Decreased GSI at downstream of Pardubice and Neratovice Decreased 11-KT levels at upstream and downstream of Neratovice | Polluted sites: Downstream and upstream of Pardubice, Neratovice, and Usti nad Labem. Pardubice receives effluents of the chemical factory containing primarily active components for making medicines and pesticides. Neratovice receives effluents of the chemical plant producing caprolactam as a raw material for polyamide fiber and the making of plastics, polyvinylchloride, and several inorganic compounds. Usti nad Labem is affected by agricultural and industrial activity Reference site: The upper reaches of the Vltava River (the tributary of the Elbe River). Concentrations (mg/kg lipid): HCHs (0.021–0.486), DDT (1.14–6.48), HCB (0.109–0.521) at polluted sites and HCHs (0.018), DDT (0.92), HCB (0.033) at reference site Sampling time: May 2004 | [140] |
| The Luvuvhu River, Limpopo Province, South Africa | Tilapia | DDT and its metabolites | Intersex at AD, ND, and XW (49%, 63%, and 58%, respectively) No intersex in the aquaria-reared fish | Polluted sites: Nandoni Dam (ND) and Xikundu Weir (XW) Reference site: Albasini Dam (AD) and reared fish | [125] |
| North West Province of South Africa | Catfish | DBP, NP, Se | Testicular abnormal growth in fish at HD during HF period. Trends toward decrease in sperm motility at HD during LF period Decreased sperm velocity at HD during both LF and HF periods | Polluted site: Hartbeespoort Dam (HD) characterized by eutrophication due to the high levels of phosphates and nitrates originating from STWs and agricultural effluent Reference site: Marico Bosveld Dam (MD) receiving water from the Marico River, an unmodified natural ecological state, Concentrations (μg/L): DBP: 3.5, NP: 4, Se 24, water chemistry: pH: 7.5 (MD) vs. 9–10.4 (HD), O2 (mg/L): 7.9 (MD) vs. 9.5–10.6 (HD), sampling time: At a low flow (LF) in Oct and at a high flow (HF) in March (MD was sampled only in Oct) | [133] |
| Lake Erie, Ontario, Canada | Goldfish | PCB | No difference in GSI Decreased T and 11-KT levels Decreased Vtg levels | Polluted site: Wheatley Harbour, Northwest shore of Lake Erie Reference site: Hillman Marsh | [128] |
| The Grand River Watershed in Southern Ontario, Canada | Rainbow darter | MWWE | Sampling time: September 2010 Decreased GSI at DS2 and DS3.5 compared to US1 Intersex at DS2 and DS3.5 compared to US1 Increased Vtg mRNA levels at DS2 compared to US1 and DS3.5 Sampling time: April and May 2011 Intersex (%): DS1 (27.3), INT1 (48.5), DS2 (84.6), DS2x (80), DS3.5 (53.3), DS Gp (6.7) vs. US1 (6.3), US2 (0), US Gp (25) No difference in GSI of normal males among US1, US2, DS1, INT1, DS2, DS2x, and DS3.5 Higher GSI of normal males at DS Gp than UP Gp No difference in GSI of intersex males No difference in E2 levels Decreased T levels at DS1, DS2, and DS3.5 Decreased 11-KT levels at DS1, INT1, DS2, and DS3.5 Decreased T and 11-KT levels in intersex males Decreased proportion of spermatogonia at DS2 Decreased proportion of spermatocytes and increased spermatids at INT1, DS2x, and DS3.5, no difference in proportion of sperm | Polluted site: Downstream Waterloo treatment plant outfall (DS1), between Waterloo and Kitchener MWWTPs (INT1), downstream Kitchener treatment plant outfall (DS2), downstream Kitchener outfall out of the plum. (DS2x), far downstream Kitchener treatment plant outfall (DS3.5) and downstream Guelph treatment plant outfall (DS Gp) Reference sites: Upstream urban areas (US1), upstream Waterloo outfall (US2) and upstream Guelph treatment plant outfall (US Gp) Sampling time: September 2010, April and May 2011 | [129,130] |
| The Alibori River, Benin, West Africa | African catfish | o,p′-p,p′-DDT, o,p′-p,p′-DDE, o,p′-p,p′-DDD, endosulfan, Heptachlor | Intersex (25–50%) Decreased 11-KT levels at S3-RS, S4-RS, S3-DS Increased E2 levels at S2-RS, S3-RS and S4-RS, S4-DS Decreased spermatocytes, spermatids, and sperm in the testes of fish from S2-S4 compared to S1 | Polluted sites: The Alibori River in Sori, Gogounou (S2), Alibori K, Kandi (S3), and Batran, Banikoara (S4). Alibori River collects the drainage from agricultural areas in the cotton-producing basin Reference site: The Pendjari River within the Pendjari National Park (S1). Agriculture is strictly forbidden Concentrations (μg/L): p,p′-DDT (0.11 RS, 0.07 DS, S4), o,p′-DDE (0.17, RS-S2, 0.19 RS-S3, 0.09 RS-S4, 0.18 DS-S2, 0.14 DS-S3, 0.17 DS-S4), p,p′-DDE, (0.13 RS-S2, 0.10 RS-S3, 0.18 RS-S4, 0.07 DS-S2, 0.08 DS-S3, 0.14 DS-S4), o,p′-DDD (0.07 RS-S2, 0.09 RS-S3, 0.09 RS-S4), p,p′-DDD (0.09 RS-S2, 0.08 RS-S4, 0.08 DS-S4), endosulfan (8.2 RS-S2, 8.8 RS-S3, 13.8 RS-S4, 2.9 DS-S2, 3.4 DS-S3, 3.7 DS-S4), Heptachlor (0.5 RS-S2, 0.5 RS-S3, 0.6 RS-S4, 1.4 DS-S2, 1.5 DS-S3, 1.3 DS-S4) Sampling time: Rainy season (RS), Sep 2010 and 2011, period of flooding with intense use of pesticides in cotton fields; dry season (DS), Feb 2011 and 2012, period of no agriculture but limited use of pesticides | [136] |
| Guinean tilapia | Intersex (12.4-40%) Decreased 11-KT levels at S2-RS, S4-RS, and S4DS Increased E2 levels at S2, S3, and S4 both RS and DS Decreased spermatids and sperm in the testes of fish from S2–S4 compared to S1 | ||||
| Grand River Watershed, Southern Ontario, Canada | Rainbow darter | MWWE | Decreased GSI at DSK and DSK2 Higher intersex at DSK, DSK2, and USK Decreased 11-KT levels at DSK, DSK2, DSW, USK, and USWDecreased T levels at DSK Increased sperm volume at DSK and DSK2 Decreased fertilization rate at DSK and USK Decreased hatching rate at DSK, USK, DSW, and USW | Polluted sites: Sites 1 km downstream of the Kitchener MWWTP (DSK) and Waterloo MWWTP (DSW), and 3 km downstream of Kitchener MWWTP (DSK2) Reference site: Sites 1 km upstream of the Kitchener MWWTP (USK) and Waterloo MWWTP (USW), and a site upstream of the city-limits (R) Sampling time: Apr and May 2012, May 2013 | [134] |
| Lake Mead National Recreation Area, USA | Carp | DDT PCB PCP PBDEs | Decreased GSI at BB, GB, LVB and WB Decreased 11-KT levels at BB, GB, LVB, and WB Decreased E2 levels at BB, GB, and WB Decreased sperm motility at LVB and WB Decreased sperm viability at LVB Decreased sperm mitochondrial functions at LVB Increased sperm chromosome breakage at LVB | Polluted sites: Boulder Basin (BB), Gregg Basin (GB), Las Vegas Bay (LVB) and Willow beach (WB), Reference site: Overton Arm (OA) DDT (ng/g wet weight): 43.4 (BB), 10.5 (GB), 107.5 (LVB), 49.7 (WB), 21.3 (OA) PCBs (ng/g wet weight): 56 (BB), 14 (GB), 94 (LVB), 381 (WB), 16 (OA) PCP (ng/g wet weight): 11 (BB), 0 (GB), 82 (LVB), 0 (WB), 0 (OA) PBDEs (ng/g wet weight): 62.9 (BB), 6.4 (GB), 118.8 (LVB), 88.4 (WB), 21.1 (OA) Sampling time: May 1998, May 1999–May 2000, March–April 2006 | [137] |
6. Laboratory Evidences for Environmental Contaminants (ECs)-Related Fertility Threat in Male Fishes
6.1. Effects of Environmental Contaminants (ECs) on Fertility
| ECs | Fish model | Dose (µg/L) | Exposure | Fertility endpoints (%) | Description | Authors | |
|---|---|---|---|---|---|---|---|
| Fertilization Rate | Hatching Rate | ||||||
| E2 | Rainbow trout | Ctrl | Adult, 50 d | 62 | Sperm were prediluted in MIS1 and added into eggs at ratio 55,000:1 to 65,000:1. Fertilizing medium was water at 6 mL. The eyed eggs were recoded 30–35 d post fertilization. | [156] A | |
| 0.0004 | 59 | ||||||
| 0.0011 | 21 * | ||||||
| 0.0026 | 7 * | ||||||
| Medaka | Ctrl | Adult, 25 d | 99 | 76 | Pairs were exposed for 21 d for females and for 25 d for males. During the exposure period, the spawned eggs were collected and the fertilized eggs (fertility of the mating pairs) were counted. During the last 3 d of exposure, the fertilized eggs from each pair were collected and the hatching rate was recorded. | [154] A | |
| 0.029 | 99 | 62 | |||||
| 0.056 | 90 | 71 | |||||
| 0.116 | 97 | 61 | |||||
| 0.227 | 98 | 64 | |||||
| 0.463 | 78 * | 50 | |||||
| EE2 | Fighting fish | Ctrl | Adult, 28 d | 78 | Pairs were exposed. Nests were removed and the pairs were kept in tanks for 3 d to determine fertilization rate by counting fry numbers. | [161] N | |
| 0.01 | 76 | ||||||
| 0.1 | 64 * | ||||||
| Zebrafish | Ctrl | Adult, 21 d | 64 | Males were mated with untreated females. After 14 d, the hatching rate was calculated. | [166] N | ||
| 0.025 | 6 * | ||||||
| Medaka | Ctrl | Adult, 21 d | 98 | 86 | Exposed spermiating male were kept in tanks with unexposed females for 7 d. All of the eggs were collected from each pair and fertilization and hatching rates were calculated. | [150] 3 | |
| 0.03 | 98 | 84 | |||||
| 0.06 | 86 | 61 | |||||
| 0.12 | 87 | 63 | |||||
| 0.48 | 94 | 81 | |||||
| Fathead minnow | Ctrl | Adult, 21 d | 98 | Pairs were exposed. After exposure, they were transferred to aquaria with clean water. The spawned eggs were collected and the fertilized eggs (fertility of the mating pairs) were counted 2 d after spawning. | [144] 4A | ||
| 0.0001 | 89 | ||||||
| 0.001 | 92 | ||||||
| 0.003 | 84 | ||||||
| 0.01 | 30 * | ||||||
| 0.1 | 0 * | ||||||
| Rainbow trout | Ctrl | Adult, 62 d | 60 a, 46 b, 25 c | Sperm was prediluted in MIS1 and added into eggs at ratio 300,000:1 (a), 50,000:1 (b), and 10,000:1 (c). Fertilization medium was ovarian fluid, 60 mM NaHCO3, 50 mM Tris, pH 9. The eyed eggs were recoded 28 d post fertilization | [155] A | ||
| 0.016 | 36 a,*, 23 b,*,19 c | ||||||
| 0.131 | 35 a,*, 22 b,*,17 c,* | ||||||
| Medaka | Ctrl | Adult, 21 d | 92 | Pairs were exposed and the spawned eggs were collected during the exposure period. The fertilized eggs (fertility of the mating pairs) were recorded. | [149] A | ||
| 0.033 | 95 | ||||||
| 0.064 | 93 | ||||||
| 0.116 | 92 | ||||||
| 0.261 | 91 | ||||||
| 0.488 | 83 | ||||||
| 17α-MT | Medaka | Ctrl | Adult, 21 d | 99 a, 98 b | 68 b | Pairs were exposed. During the exposure period, the spawned eggs were collected and the fertilized eggs (fertility of the mating pairs) and the hatching rate were recorded following 7 d (a) or 21 d (b) of exposure. | [145] A |
| 0.023 | 98 a, 97 b | 60 b | |||||
| 0.047 | 86 a, 80 b,* | 39 b,* | |||||
| 0.088 | 90 a, 33 b,* | 18 b,* | |||||
| 0.188 | 96 a, 0 b,* | 0 b,* | |||||
| 0.380 | 96 a, 0 b,* | 0 b,* | |||||
| Fathead minnow | Ctrl | Adult, 21 d | 98 | After exposure, 3–4 pairs were transferred to aquaria with clean water. The spawned eggs were collected and the fertilized eggs (fertility of the mating pairs) were counted 2 d after spawning. | [143] 5A | ||
| 0.1 | 98 | ||||||
| 1 | 59 | ||||||
| 5 | 31 * | ||||||
| 50 | 0 * | ||||||
| 4-NP | Medaka | 0 | Adult, 21 d | 97 | 94 | Pairs were exposed. For 21 consecutive d, spawned eggs were collected, counted, and assessed for fertilization rate. The fertilized eggs were maintained at the same treatments, and number of hatched embryos was recorded to assess hatching rate. | [167] A |
| 1.27 | 96 | 98 | |||||
| 2.95 | 97 | 96 | |||||
| 9.81 | 95 | 99 | |||||
| 27.8 | 95 | 99 | |||||
| 89.4 | 95 | 77 * | |||||
| Rainbow trout | Ctrl | Adult, 60 d | 97 a, 65 b | Sperm was prediluted in MIS1 and added into eggs at ratios of 55,000:1 to 65,000:1. Fertilizing medium was water. The eyed eggs were recoded 35 d post fertilization. Fertilization was recorded following 30 d (a) and 60 d (b) of exposure. | [146] N | ||
| 0.13 | 96 a, 64 b | ||||||
| 0.28 | 94 a, 68 b | ||||||
| 0.75 | 95 a, 0 b,* | ||||||
| Medaka | Ctrl | Adult, 21 d | 98a, 98b | Pairs were exposed. During the exposure period, the spawned eggs were collected and the fertilized eggs (fertility of the mating pairs) were counted following 7 d (a) or 21 d (b) of exposure. | [152] A | ||
| 24.8 | 98a, 96b | ||||||
| 50.9 | 95a, 95b | ||||||
| 101 | 98 a, 89 b | ||||||
| 184 | 96 a, 78 b,* | ||||||
| BPA | Rare minnow | Ctrl | Adult, 21 d | 96 a,b,c | Males were exposed to BPA for 7 d (a), 14 d (b), or 21 d (c). Fertilization was performed in vitro, and evaluated at 4 h post fertilization. | [168] A | |
| 11 | 92 a,*, 90 b,*, 89 c,* | ||||||
| 206 | 90 a,*, 89 b,*, 87 c,* | ||||||
| Zebrafish | Ctrl | Larvae, 150 d | 92 | 87 | BPA exposure was from larvae (6 d post fertilization) to 5 months post fertilization. | [164] A | |
| 0.228 | 90 | 61 * | |||||
| 2.28 | 89 | 62 * | |||||
| 22.8 | 87 * | 86 | |||||
| Zebrafish | 0.032 | Embryos, 150 d | n.s. | 47 | Four females and 4 males within the same group were maintained together to produce offspring for assessment of fertilization and hatching rates. Solvent contains 0.03 BPA. | [163] A | |
| 0.372 | n.s. | 24 * | |||||
| Brown trout | Ctrl | Adult, 76 d | 66 | Sperm of exposed males was prediluted in MIS1 and added into eggs at ratios of 45,000:1 to 60,000:1. Fertilizing medium was water. The eyed eggs were recoded 30–35 d post fertilizat | [157] N | ||
| 1.75 | 73 | ||||||
| 2.4 | 76 | ||||||
| 5.0 | 28 * | ||||||
| Medaka | Ctrl | Adult, 21 d | 99 | Pairs were exposed. During the exposure period, the spawned eggs were collected and the fertilized eggs were counted. | [147] A | ||
| 837 | 98 | ||||||
| 1720 | 99 | ||||||
| 3120 | 99 | ||||||
| BPS | Zebrafish | Ctrl | Embryos, 75 d | 90 | Embryos were exposed to BPS. Males and females were assigned to new tanks and acclimatized for 3 d. Numbers of spawned eggs were recorded daily for the next 7 d. | [162] N | |
| 0.1 | 88 | ||||||
| 1 | 85 | ||||||
| 10 | 48 * | ||||||
| 100 | 42 * | ||||||
| Ctrl | Adult, 21 d | 98 | Fertilized eggs of exposed female and male were collected at 16 d post exposure, and were exposed to same BPS concentrations until 6 d post fertilization, and hatching rate was determined. | [159] A | |||
| 0.5 | 28 * | ||||||
| 5 | 21 * | ||||||
| 50 | 10 * | ||||||
| BPAF | Zebrafish | Ctrl | Embryos, 120 d | 76 | 99 | Embryos were exposed to BPAF for 120 d. The number embryo/larvae was recorded at 7 d post fertilization. | [160] N |
| 5 | 70 | 91 | |||||
| 25 | 75 | 92 | |||||
| 125 | 49 * | 94 | |||||
| MEHP | Zebrafish | Ctrl | Pre-adult (2 month-old), 81 d | 82 a, 82 b | One hundred fertilized eggs of male and female exposed to MEHP were collected and divided into group (a) receiving the same MEHP concentrations, and group (b) receiving no further MEHP. The hatching rate was determined during 6 d post fertilization. | [165] A | |
| 0.47 | 82 a, 54 b,* | ||||||
| 4.0 | 67 a, 57 b,* | ||||||
| 37.5 | 65 a,*, 47 b,* | ||||||
| DEHP | Zebrafish | Ctrl | Adult, 21 d | 64 | Males were mated with untreated females. Over a period of 14 d, the hatching rate was calculated. | [166] N | |
| 0.2 | 3 * | ||||||
| 20 | 2 * | ||||||
| Ctrl | Adult, 10 d | 83 a, 81 b | Males were injected into the intraperitoneal cavity (mg/kg). Each aquarium contains 2 females and 2 males. Following 1–5 d (a) and 6–10 d (b) of treatment, eggs were collected and fertilization rate was recorded. | [153] A | |||
| 0.5 | 78 a, 80 b | ||||||
| 50 | 88 a, 78 b | ||||||
| 5000 | 75 a, 48 b,* | ||||||
| FLU | Medaka | Ctrl | Adult, 21 d | 95 a, 97 b, 95 c | Pairs were exposed. All spawned eggs were collected from female fish and fertility of each pair were checked daily for 3 weeks: (a) 1 week, (b) 2 weeks, and (c) 3 weeks post exposure. | [169] A | |
| 100 | 96 a, 96 b, 94 c | ||||||
| 200 | 99 a, 95 b, 99 c | ||||||
| 400 | 94 a, 100 b, 95 c | ||||||
| 790 | 96 a, 99 b, 95 c | ||||||
| 1560 | 18 a,*, 50 b,*, 49 c,* | ||||||
| VZ | Fathead minnow | 0, 60, 250. 450 | Adult, 28 d | n.s. | n.s. | Pairs were exposed. The spawned eggs were collected and the fertilized eggs (fertility of the mating pairs) and hatching rate were counted during exposure. | [148] A |
| MCP | Guppy | Ctrl | Embryos, 90 d | 34 | Pairs of guppies were exposed in a semi-static exposure system. Number of newly hatched offspring produced per female were counted. | [158] N | |
| 10 | 21 * | ||||||
| 100 | 10 * | ||||||
| 1000 | 9 * | ||||||
| MMS | Brown trout | Ctrl | Adult, 21 d | 94 | 93 | Males were injected into the intraperitoneal cavity (mg/kg). Sperm (100 μL) were added into 40 g eggs. Fertilization medium was SAFD2. Fertilization rate was checked after 120 degree-days of development of eggs. | [151] A |
| 50 | 95 | 94 | |||||
| Arctic charr | 0 | Adult, 21 d | 84 | 75 | |||
| 50 | 83 | 76 | |||||
6.2. Effects of Environmental Contaminants (ECs) on Sperm Production
| ECs | Model | Dose (µg/L) | Exposure | Spermiation (%) | Volume (mL) | Density (×109 sperm/mL) | Authors and Notes |
|---|---|---|---|---|---|---|---|
| E2 | Grayling | Ctrl | Adult, 50 d | 0.3 | Authors: [156] | ||
| 0.0011 | 0.1 * | ||||||
| Rainbow trout | Ctrl | Adult, 50 d | 4.1 a, 3.8 b | 6.1 a, 6.3 b | Authors: [156]; Analysis performed 35 d (a) and 50 d (b) of exposure; Sperm volume: Sperm mass (g) | ||
| 0.0004 | 4.0 a, 2.8 b | 6.0 a, 6.5 b | |||||
| 0.0011 | 2.0 a,*, 1.5b,* | 5.6 a, 5.4 b,* | |||||
| 0.0026 | 2.2 a, 1.4 b,* | 5.7 a, 3.9 b,* | |||||
| Goldfish | Ctrl | Adult, 70 d | 91 a, 94 b | Authors: [170]; This study was performed in winter (a) and summer (b). | |||
| 0.05 | 67 a,*, 40 b,* | decrease (data not shown) | decrease (data not shown) | ||||
| EE2 | One-sided livebearer | Ctrl | Adult, 28 d | 2.4 | Authors: [182]; Sperm density: ×106 sperm/mL. | ||
| 0.005 | 2.2 | ||||||
| 0.064 | 2.2 | ||||||
| 0.127 | 2.0 | ||||||
| Rainbow trout | Ctrl | Adult, 62 d | 7.9 | Authors: [155] | |||
| 0.016 | 20 * | ||||||
| 0.131 | 30 * | ||||||
| 4-NP | Rainbow trout | Ctrl | Adults, 60 d | 5.9 a, 6.0 b | 7.2 a, 7.0 b | Authors: [146]; Analysis performed at 30 d (a) and 60 d (b) of exposure | |
| 0.13 | 3.8 a,*, 1.5 b,* | 7.4 a, 6.8 b | |||||
| 0.28 | 4.0 a,*, 1.0 b,* | 6.9 a, 6.6 b | |||||
| 0.75 | 2.7 a,*, 0.1 b,* | 7.3 a, n.d. b,* | |||||
| BPA | Rare minnow | Ctrl | Adult, 21 d | 2.0 a, 2.0 b, 1.9 c | Authors: [168]; Analysis performed at 7 d (a), 14 d (b), and 21 d (c) of exposure; Sperm density: ×1013 sperm/male | ||
| 11 | 2.0 a, 2.1 b, 1.9 c | ||||||
| 206 | 2.2 a, 1.9 b, 1.8 c | ||||||
| Zebrafish | Ctrl | Embryos, 150 d | 1.2 | 4.0 | Authors: [164]; Sperm volume: ×107 sperm/testis; Sperm density: ×109 sperm/g testis | ||
| 0.228 | 1.0 * | 4.0 | |||||
| 2.28 | 1.1 | 4.0 | |||||
| 22.8 | 1.2 | 4.0 | |||||
| Ctrl | Larvae, 150 d | 1.3 | 4.5 | ||||
| 0.228 | 0.8 * | 3.5 * | |||||
| 2.28 | 1.2 | 4.5 | |||||
| 22.8 | 1.3 | 4.5 | |||||
| Ctrl | Adult, 150 d | 1.2 | 3.5 | ||||
| 0.228 | 1.1 | 3.5 | |||||
| 2.28 | 1.2 | 3.5 | |||||
| 22.8 | 0.6 * | 1.8 * | |||||
| Zebrafish | Solvent | Embryos, 150 d | 2.8 | Authors: [163]; Solvent contains 0.03 BPA. Sperm density: ×109 sperm/g testis | |||
| 0.37 | 2.2 * | ||||||
| Goldfish | Ctrl | Adult, 90 d | 207.1 | 12.9 | Authors: [173,174]; Sperm volume: μL | ||
| 0.2 | 52.9 * | 11.1 | |||||
| 2.0 20 | 73.3 * 58.8 * | 10.8 9.2 * | |||||
| Brown trout | Ctrl | Adult, 76 d | 70 a, 80 b, 70c | 0.5 a, 0.5 b, 0.5 c | 29.1 a, 25.1 b, 23.3c | Authors: [157]; Analysis performed at day 38 (a), 56 (b), and 76 (c) of exposure; Sperm volume: Sperm mass (g) | |
| 1.75 | 80 a, 80 b, 60c | 0.6 a, 0.6 b, 0.5 c | 26.6 a,*, 26.0 b, 24.1 c | ||||
| 2.4 | 60 a, 80 b, 60c | 0.6 a, 0.5 b, 0.7 c | 26.8 a,*, 25.8 b, 24.2 c | ||||
| 5 | 10 a, b, c,و | 0.1 a,*, 0.02 b,*, 0.1 c | 26.2 a,*, 25.2 b, 25.4 c | ||||
| Guppy | Ctrl | Adult, 21 d | 8.0 | Authors: [172]; Sperm density: ×106 sperm/mL | |||
| 274 | 5.1 * | ||||||
| 549 | 2.0 * | ||||||
| BPS | Zebrafish | Ctrl | Embryos, 75 d | 7.8 | Authors: [162] | ||
| 0.1 | 7.1 | ||||||
| 1 | 6.9 | ||||||
| 10 | 4.5 * | ||||||
| 100 | 2.9 * | ||||||
| FLX | Mosquitofish | Ctrl | Adult, 30 d | 13 | Authors: [183]; Sperm density: ×106 sperm/g testis | ||
| 0.042 | 19 * | ||||||
| 0.479 | 17 * | ||||||
| Goldfish | Ctrl | Adult, 14 d | 23 a, 0.9 b | Authors: [181]; Analysis performed at 7 d (a) and 14 d (b) of exposure; Sperm volume: μL | |||
| 0.375 | 10 a,*, 0.5 b | ||||||
| 45 | 0.7 a,*,0.2 b,* | ||||||
| VZ | Goldfish | Ctrl | Adult, 30 d | 90 | Authors: [178]; Sperm volume: μL | ||
| 100 | 53 * | ||||||
| 400 | 15 * | ||||||
| 800 | 6 * | ||||||
| Guppy | Ctrl | Adult, 30 d | 3.0 | Authors: [177]; VZ: μg per mg dry food; Sperm density: ×106 sperm/male | |||
| 0.1 | 2.3 | ||||||
| 1 | 1.7 * | ||||||
| 10 | 1.0 * | ||||||
| Guppy | Ctrl | Adult, 30 d | 4.9 | Authors: [176]; VZ: μg per mg dry food; Sperm density: ×106 sperm/male | |||
| 1 | 3.6 | ||||||
| 10 | 3.1 | ||||||
| MEHP | Zebrafish | Ctrl | Pre-adult (2 month-old), 81 d | 8.3 | 9.8 | Authors: [165]; Sperm volume: ×107 sperm/d; Sperm density: ×1010 sperm/mL | |
| 0.46 | 7.4 | 9.1 | |||||
| 4.0 | 7.1 | 8.6 * | |||||
| 37.5 | 6.8 * | 7.2 | |||||
| DEHP | Goldfish | Ctrl | Adult, 30 d | 90.4 | Authors: [175]; Sperm volume: μL | ||
| 1 | 3.7 * | ||||||
| 10 | 40.8 * | ||||||
| 100 | 8.7 * | ||||||
| FLU | Zebrafish | Ctrl | Adult, 30 d | 1.5 | Authors: [179]; Sperm density: ×107 sperm/mL | ||
| 0.279 | 0.7 * | ||||||
| Guppy | Ctrl | Adult, 30 d | 4.9 | Authors: [176]; Flu: μg Flu per mg dry food Sperm density: ×106 sperm/male | |||
| 1 | 2.9 * | ||||||
| 10 | 1.4 * | ||||||
| DES | Zebrafish | Ctrl | Adult, 30 d | 1.5 | Authors: [179]; Sperm density: ×107 sperm/mL | ||
| 0.095 | 0.5 * | ||||||
| TBT | Guppy | Ctrl | Adult, 21 d | 8.0 | Authors: 170; Sperm density: ×106 sperm/mL | ||
| 0.011 | 3.0 * | ||||||
| 0.022 | 2.7 * | ||||||
| Zebrafish | Ctrl | Embryos, 70 d | 0.5 a, 0.7 b | Authors: [171]; Analysis performed at day 30 (a) and 70 (b) of exposure. At 0.01 µg/L no male was spermiated. | |||
| 0.00001 | 0.4 a, 1.1 b | ||||||
| 0.001 | 0.6 a, 2.1 b,* | ||||||
| 0.1 | 1.9 a,*, 3.0 b,* | ||||||
| p,p ′-DDE | Guppy | Ctrl | Adult, 30 d | 4.9 | Authors: [176]; p,p′-DDE: μg per mg dry food; Sperm density: ×106 sperm/male | ||
| 0.1 | 3.8 * | ||||||
| 1 | 6.7 * | ||||||
| MCP | Guppy | Ctrl | Embryos, 90 d | 3.4 | Authors: [158]; Sperm density: ×106 sperm/mL | ||
| 10 | 1.9 * | ||||||
| 100 | 2.2 * | ||||||
| 1000 | 2.0 * | ||||||
| Clofibrate | Fathead minnow | Ctrl | Adult, 21 d | 4.8 a, 1.2 b, 1.0 c | Authors: [180]; a, 53–57 females: 8–12 males; b, 6 females: 6 males, and c, 2 females: 10 males; Sperm volume is measured in ×106 sperm/mg testis | ||
| 1 | 0.8 c | ||||||
| 10 | 0.3 b,*, 0.8 c | ||||||
| 1000 | 1.7 a,*, 0.5 b,*, 1.1 c | ||||||
| MSTPs | Goldfish | Ctrl | Adult, 70 d | 91a, 94b | Authors: [170]; This study was performed in winter (a) and summer (b) | ||
| 100% | 100a, 93b | n.s. | n.s. |
6.3. Effects of Environmental Contaminants (ECs) on Sperm Morphology
6.4. Effects of Environmental Contaminants (ECs) on Sperm Genome
6.5. Effects of Environmental Contaminants (ECs) on Sperm Motility Kinetic
| ECs | Model | Dose (µg/L) | Exposure | TPA (s) | Motility (%) | Velocity (µm/s) | Authors and Notes | ||
|---|---|---|---|---|---|---|---|---|---|
| VCL | VSL | VAP | |||||||
| E2 | One-sided livebearer | Ctrl | Adult, 28 d | 240 (every 10 s) | 79 | 113 | 110 | Authors: [197]; AM: HAMF-10 medium | |
| 0.05 | 78 | 115 | 111 | ||||||
| 0.10 | 70 | 115 | 111 | ||||||
| 0.25 | 75 | 118 | 116 | ||||||
| Rainbow trout | Ctrl | Adult, 50 d | 10 | 89 | 78 | Authors: [156]; IM (mM): 103 NaCl, 40 KCl, 1 CaCl2, 0.8 MgSO2, 20 Tris, pH 7.8; AM (mM): 60 NaHCO3, 20 Glycin, pH 9.0; Analysis done at 35 d post exposure. | |||
| 0.0004 | 87 | 82 | |||||||
| 0.0011 | 85 | 84 | |||||||
| 0.0026 | 80 | 90 | |||||||
| Grayling | Ctrl | Adult, 50 d | 10 | 74 | 105 | Authors: [156]; IM and AM: See above row | |||
| 0.001 | 46 * | 78 * | |||||||
| Goldfish | Ctrl | Adult, 70 d | n.d. | 2500 a, 3000 b | 100 a, 62 b | Authors: [170]; IM: Ringer solution; AM: DW; Motility was absolute number of motile sperm per fish. Exposures were performed in winter (a) and summer (b). | |||
| 0.05 | <10 a,*, <10 b,* | 112 a, 75 b | |||||||
| P | Fathead minnow | Ctrl | Adult, 7 d | 80 (every 15 s) | 53 | 94 | 75 | 90 | Authors: [194]; IM: 0.8 mM NaCl solution. AM: DW |
| 0.025 | 63.5 | 78 | 52 | 73 | |||||
| 0.339 | 39 | 60 * | 38 * | 53 * | |||||
| EE2 | Zebrafish | Ctrl | Embryos, 240 d | 44 (every 2 s) | 58 | 89 | 56 | Authors: [198]; IM: n.d.; AM: n.d.; Solvent: DMSO | |
| Solvent | 49 | 80 | 47 | ||||||
| 0.0002 | 53 | 87 | 49 | ||||||
| 0.00024 | 38 | 79 | 49 | ||||||
| 0.001 | 47 | 88 | 52 | ||||||
| one-sided livebearer | Ctrl | Adult, 28 d | 240 (every 10 s) | 82 | 103 | 101 | Authors: [182]; AM: HAMF-10 medium | ||
| 0.005 | 75 | 109 | 105 | ||||||
| 0.064 | 76 | 107 | 103 | ||||||
| 0.127 | 65 * | 106 | 109 | ||||||
| Fighting fish | Ctrl | Adult, 28 d | 60 (every 6 s) | Authors: [161]; IM (g/L): 5.5 NaCl, 2 KCl, 3.8 Glycine, 2.4 Tris, pH 7.5; AM: Water containing EE2 | |||||
| 0.01 | n.s. | n.s. | n.s. | n.s. | |||||
| 0.1 | n.s. | n.s. | decrease | n.s. | |||||
| Medaka | Ctrl | Adult, 21 d | 30 | 70 | 60 | Authors: [150]; AM (g/L): 1 NaCl, 0.03 KCl, 0.04 CaCl2, 0.1 MgCl2, 0.2 NaHCO3, pH 7.3 | |||
| 0.06 | 88 | 85 | |||||||
| 0.12 | 85 | 83 | |||||||
| 0.24 | 90 | 75 | |||||||
| 0.48 | 83 | 81 | |||||||
| Rainbow trout | Ctrl | Adult, 62 d | 12 (every 15 s) | Authors: [155]; IM (mM): 103 NaCl, 40 KCl, 1 CaCl2, 0.8 MgSO2, 20 Tris, pH 7.8; AM (mM): 60 NaHCO3, 50 Tris, pH 9 | |||||
| 0.016 | n.s. | n.s. | n.s. | ||||||
| 0.131 | n.s. | n.s. | n.s. | ||||||
| LNG | Fathead minnow | 0 | Adult, 14 d | 80 (every 15 s) | 85 | 68 | 81 | 52 | Authors: [192] |
| 0.012 | 74 * | 60 | 62 * | 48 | |||||
| 0.127 | 45 * | 47* | 58 * | 46 | |||||
| BPA | Rare minnow | Ctrl | Adult, 21 d | n.d. | 97 a, b, c | Authors: [168]; IM: Hank’s balance; AM: 0.55% NaCl | |||
| 11 | 97 a, 97 b, 96 c | ||||||||
| 206 | 97 a, 96 b, 95 c | ||||||||
| Zebrafish | Ctrl | Embryos, 150 d | n.d. (15 frames each 0.05 s) | 65 | 128 | 74 | 87 | Authors: [164]; IM: Hank’s balance; AM: n.d. | |
| 0.228 | 53* | 120 * | 58 * | 73* | |||||
| 2.28 | 61 | 113 * | 50 * | 68* | |||||
| 22.8 | 61 | 116 | 62 | 76 | |||||
| Ctrl | Larvae, 150 d | 68 | 128 | 66 | 86 | ||||
| 0.228 | 55 * | 128 | 66 | 84 | |||||
| 2.28 | 61 | 117 * | 62 | 76 | |||||
| 22.8 | 62 | 120 | 58 * | 72 * | |||||
| Ctrl | Adult, 150 d | 65 | 130 | 74 | 87 | ||||
| 0.228 | 60 | 125 * | 68 | 84 | |||||
| 2.28 | 59 | 125 * | 62 * | 76 * | |||||
| 22.8 | 45 * | 125 * | 60 * | 76 * | |||||
| Zebrafish | Solvent | Embryos, 150 d | n.d. (15 frames each 0.05 s) | 75 | 100 | Authors: [163]; IM: Hank’s balance; AM: n.d. Solvent contains 0.03 BPA | |||
| 0.37 | 36 * | 68 | |||||||
| Goldfish | Ctrl | Adult, 30 d | 15 | 97 a, 100 b, 99 c | 157 a, 146 b, 166 c | Authors: [173];AM (mM): 50 NaCl, 20 Tris, pH 8.5, osmolality 110 mOsmol/kg; Analyses performed at day 10 (a), 20 (b), and 30 (c) of exposure | |||
| 0.6 | 94 a, 94 b, 95 c | 148 a, 156 b, 159 c | |||||||
| 4.5 | 96 a,80 b,*,88 c,* | 158 a, 136 b, 158 c | |||||||
| 11.0 | 98 a, 85 b,*,91 c,* | 144 a,138 b, 154 c | |||||||
| Ctrl | 60 | 90 a, 79 b, | 55 a, 30 b, 38 c | ||||||
| 0.6 | 78 a, 74 b, 70 c,* | 34 a,*, 25 b,*, 30 c,* | |||||||
| 4.5 | 85a, 61b,*, 68c,* | 29 a,*, 34 b,*, 38 c,* | |||||||
| 11.0 | 85 a, 65 b,*, 61 c,* | 30 a,*, 29 b,*, 37 c,* | |||||||
| Goldfish | Ctrl | Adult, 90 d | 15 | 99 | 168 | Authors: [174]; AM (mM): 50 NaCl, 5 KCl, 20 Tris, pH 8.5, osmolality 110 mOsmol/kg | |||
| 0.2 | 94 | 155 | |||||||
| 2 | 95 | 150 * | |||||||
| 20 | 88* | 140 * | |||||||
| Ctrl | 60 | 64 | 60 | ||||||
| 0.2 | 51 * | 59 | |||||||
| 2 | 46 * | 48 | |||||||
| 20 | 41 * | 47 * | |||||||
| Brown trout | Ctrl | Adult, 76 d | 10 | 84 a, 88 b, 86 c | 97 a, 107 b, 106 c | Authors: [157]; IM (mM): 103 NaCl, 40 KCl, 1 CaCl2, 0.8 MgSO2, 20 Tris, pH 7.8; AM (mM): 60 NaHCO3, 20 Glycin, pH 9.0; Analyses were performed at day 38 (a), 56 (b), and 76 (c) of exposure | |||
| 1.75 | 33 a,*, 73 b, 70 c | 62 a,*, 75 b,*, 111 c | |||||||
| 2.4 | 40 a,*, 36 b,*, 81 c | 75 a,*, 75 b,*, 92 c | |||||||
| 5 | 4 a,*, 1 b,*, 4 c | 32 a,*, 49 b,*, 41 c,* | |||||||
| 4-NP | Rainbow trout | Ctrl | Adult, 60 d | 10 | 68 a, 75 b, 69 c | 82 a, 92 b, 89 c | Authors: [146]; IM (mM): 103 NaCl, 40 KCl, 1 CaCl2, 0.8 MgSO2, 20 Tris, pH 7.8; AM (mM): 60 NaHCO3, 20 Glycin, pH 9.0; Analyses were performed at day 0 (a), 30 (b), and 60 (c) of exposure | ||
| 0.13 | 69 a, 70 b, 65 c | 77 a, 96 b, 84 c | |||||||
| 0.28 | 72a, 75b, 81c | 81a, 101b, 78c | |||||||
| 0.75 | 66a, 70b, n.d.c,* | 82a, 91b, n.d.c,* | |||||||
| FLX | Mosquitofish | Ctrl | Adult, 35 d | 85 | 112 | 62 | 80 | Authors: [196]; IM (mM): 207 NaCl, 5.4 KCl, 1.3 CaCl2, 0.49 MgCl2, 0.41 MgSO4, 10 Tris, pH 7.5; AM: 150 mM KCl with 2 mg/mL BSA | |
| 0.031 | 83 | 115 | 65 | 84 | |||||
| 0.375 | 84 | 108 | 61 | 77 | |||||
| VZ | Goldfish | Ctrl | Adult, 30 d | 15 | 99 | 165 | Authors: [178]; AM (mM): 50 NaCl, 20 Tris, pH 8.5 Osmolality 110 mOsmol/kg | ||
| 100 | 97 | 159 | |||||||
| 400 | 80 | 137 | |||||||
| 800 | 19* | 130* | |||||||
| Ctrl | 60 | 64 | 64 | ||||||
| 100 | 52 | 58 | |||||||
| 400 | 44* | 65 | |||||||
| 800 | 14* | 60 | |||||||
| DEHP | Goldfish | Ctrl | Adult, 30 d | 15 | 98 | 161 | Authors: [175]; AM (mM): 50 NaCl, 5 mM KCl. 20 Tris, pH 8.5 | ||
| 1 | 86 | 142 | |||||||
| 10 | 88 | 128* | |||||||
| 100 | 74* | 107* | |||||||
| Ctrl | 60 | 67 | 67 | ||||||
| 1 | 46* | 60 | |||||||
| 10 | 47* | 63 | |||||||
| 100 | 38* | 68 | |||||||
| Clofibrate | Fathead minnow | Ctrl | Adult, 21 d | 5 | 62 | 30 | 22 | Authors: [180]; IM (mM): 94 NaCl, 27 KCl, 50 Glycine, 15 Tris pH. 7.6; AM: Water; Sperm velocity in nm/s. | |
| 1 | 56 | 26 | 16 | ||||||
| 10 | 51 | 24 | 18 | ||||||
| 1000 | 50 | 21 * | 11 * | ||||||
| TBT | Zebrafish | Ctrl | Fry, 70 d | 5 | 78 a, 60 b | Authors: [171]; IM (g/L): 5.8 NaCl, 0.2 KCl, 0.2 CaCl2, 0.04 MgCl2, 2.1 NaHCO3, 0.04 NaH2PO4, 3.8 glycine, pH 8.6; AM: DW; Analyses were performed at day 30 (a) and 70 (b) of exposure | |||
| 0.00001 | 62 a, 62b | ||||||||
| 0.001 | 44 a,*, 19 b,* | ||||||||
| 0.01 | n.d. a,*, 20 b,* | ||||||||
| 0.1 | 36 a,*, 0 b,* | ||||||||
| MSTPs | Goldfish | Ctrl | Adult, 70 | n.d. | 2500 a, 3000 b | 100 a, 62 b | Authors: [170]; IM: Ringer solution; AM: DW; Sperm motility shows absolute number of motile sperm per fish. Exposures were performed in winter (a) and summer (b) | ||
| 100% | 2000 a, 2700 b | 100 a, 95 b | |||||||
| Cu | Killifish | Ctrl | Fry, 345 d | 83 | Authors: [193]; Ctrl contains 1.8 Cu | ||||
| 7.1 | 26 * | ||||||||
| 10.9 | 17 * | ||||||||
| Alderin | Catfish | 0 | 75 | 109 | 89 | 103 | Authors: [195]; IM (g): 5.49 NaCl, 2.01 KCl, 3.75 glycine, 1.82 Tris; AM: Tank water | ||
| 0.14 | 71 | 100 | 80 | 91 | |||||
| DMDT | 0.23 | 68 | 86 | 71 | 81 | ||||
| ECs | Environmental Concentration (ng/L) | Fish Species | LTC (ng/L) | HTC (ng/L) | Fertility | Sperm | Authors | ||
|---|---|---|---|---|---|---|---|---|---|
| Production | Motility | Velocity | |||||||
| E2 | 0.1–5.01, 0.2–2.92, 0.63, 0.5–5.24, 0.4–3.35, 0.3–55.06, 2.7–48.07, LOD-3.711, 7.4820, LOD-7.426, LOD-84.327, LOD-33.428, <1–17529 | Zebrafish | - | 25 | + | n.d. | n.d. | n.d. | [166] N |
| One-sided livebearer | 50 | 250 | n.d. | n.d. | - | - | [197] A | ||
| Rainbow trout | 0.4 | 2.6 | + | + | - | - | [156] N | ||
| Medaka | 29 | 463 | - | n.d. | n.d. | n.d. | [154] A | ||
| Grayling | - | 1.0 | n.d. | + | + | + | [156] N | ||
| Goldfish | - | 50 | n.d. | + | + | - | [170] A | ||
| P | 7.4–11.81, 5.4–6.18 | Fathead minnow | 25 | 339 | n.d. | n.d. | - | - | [194] A |
| EE2 | 0.7–<2.02, 0.1–8.94, 0.045, 0.2–7.59, LOD-42.010, LOD-0.811; LOD-35.628, <0.8–34.029 | Fighting fish | 10 | 100 | - | n.d. | - | - | [161] N |
| Zebrafish | 0.2 | 1 | n.d. | n.d. | - | - | [198] A | ||
| Medaka | 30 | 480 | - | n.d. | n.d. | - | [150] A | ||
| Fathead minnow | 0.1 | 100 | + | n.d. | n.d. | n.d. | [144] 1 | ||
| Rainbow trout | 16 | 131 | + | + | - | - | [155] A | ||
| Medaka | 33 | 488 | - | n.d. | n.d. | n.d. | [149] A | ||
| One-sided livebearer | 10 | 150 | n.d. | - | - | - | [182] A | ||
| 17α-MT | 1.3–1.88, <0.9–14.512, 1.3313 | Medaka | 23 | 380 | - | n.d. | n.d. | n.d. | [145] A |
| Fathead minnow | 100 | 50,000 | - | n.d. | n.d. | n.d. | [143] 2 | ||
| 4-NP | LOD-37,00011, LOD-10,18614, 15–38615, 77–114216, 112.4–2065.726, <0.5–211.029 | Medaka | 1,270 | 89,400 | - | n.d. | n.d. | n.d. | [167] A |
| Rainbow trout | 130 | 750 | + | + | + | + | [146] N | ||
| Medaka | 248 | 184,000 | - | n.d. | n.d. | n.d. | [152] A | ||
| BPA | LOD-12,20514, LOD-112516, LOD-800017, 0.5–70218, 6.6–74.919, 12.3–755.626, <1–14529 | Rare minnow | 11,000 | 206,000 | + | - | - | n.d. | [168] A |
| Zebrafish | 228 | 22,800 | + | + | + | + | [163,164] A | ||
| Brown trout | 1,750 | 5,000 | + | + | + | + | [157] N | ||
| Medaka | 837,000 | 3,120,000 | - | n.d. | n.d. | n.d. | [147] A | ||
| Goldfish | 200 | 20,000 | n.d. | + | + | + | [173] A, [174] N | ||
| Guppy | 274,000 | 549,000 | n.d. | - | n.d. | n.d. | [172] N | ||
| BPS | 0.3–19.019 | Zebrafish | 100 | 100,000 | - | - | n.d. | n.d. | [162] N |
| Zebrafish | 500 | 50,000 | - | + | n.d. | n.d. | [159] A | ||
| BPAF | LOD-15,00043 | Zebrafish | 5000 | 125,000 | - | n.d. | n.d. | n.d. | [160] N |
| DEHP | 330–182,00018, 230–73020, 364–2.6821, 61.6–4352.022, 150–12,10041 | Zebrafish | 200 | 20,000 | + | n.d. | n.d. | n.d. | [166] N |
| Zebrafish | 500 | 5,000,000 | - | n.d. | n.d. | n.d. | [153] A | ||
| Goldfish | 1000 | 100,000 | n.d. | + | + | + | [175] N | ||
| VZ | 0.5–2023, 524, <1025 | Fathead minnow | 60,000 | 450,000 | - | n.d. | n.d. | n.d. | [148] A |
| Goldfish | 100,000 | 800,000 | n.d. | - | - | - | [178] N | ||
| Guppy | 100 | 10,000 | n.d. | - | n.d. | n.d. | [177] A | ||
| Guppy | 1000 | 10,000 | n.d. | - | n.d. | n.d. | [176] A | ||
| DES | LOD-3.327, LOD-8.528 | Zebrafish | - | 95 | n.d. | - | n.d. | n.d. | [179] A |
| FLX | 0.4–2.630, 2.0–19.531; 4.7–9.432; LOD-12833 | Mosquitofish | 42 | 479 | n.d. | + | - | - | [183] A, [197] A |
| Goldfish | 375 | 45,000 | n.d. | ± | n.d. | n.d. | [181] A | ||
| FLU | 0.55–1.134, 12–3044 | Zebrafish | - | 279 | n.d. | ± | n.d. | n.d. | [179] A |
| Medaka | 100,000 | 1,560,000 | - | [169] A | |||||
| Guppy | 1000 | 10,000 | n.d. | ± | n.d. | n.d. | [176] A | ||
| Clofibrate | LOD-17.232; 0.2–0.735, 6–700044 | Fathead minnow | 1000 | 1,000,000 | n.d. | ± | - | - | [180] A |
| MCP | 8.336, 16537, LOD-400038 | Guppy | 10,000 | 1,000,000 | ± | ± | n.d. | n.d. | [158] N |
| MMS | 0.139 | Brown trout | - | 50,000 | - | - | n.d. | n.d. | [151] N |
| Arctic charr | - | 50,000 | - | - | n.d. | n.d. | [151] N | ||
| TBT | 0.4240 | Guppy | 11 | 22 | n.d. | ± | n.d. | n.d. | [172] N |
| Zebrafish | 0.01 | 100 | n.d. | ± | n.d. | ± | [171] N | ||
| DDE | 36042 | Guppy | 100 | 1000 | n.d. | + | n.d. | n.d. | [176] A |
| Aldrin | 14042 | African catfish | 140 | n.d. | n.d. | - | - | [196] N | |
| DMDT | 23042 | African catfish | 230 | n.d. | n.d. | - | - | [196] N | |
7. Conclusions
8. Future Research Directions
9. Suggestions to Normalize Future Studies
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Richiardi, L.; Bellocco, R.; Adami, H.O.; Torrang, A.; Barlow, L.; Hakulinen, T.; Rahu, M.; Stengrevics, A.; Storm, H.; Tretli, S.; et al. Testicular cancer incidence in eight Northern European countries: Secular and recent trends. Cancer Epidemiol. Prev. Biomark. 2004, 13, 2157–2166. [Google Scholar]
- Andersson, A.M.; Jørgensen, N.; Main, K.M.; Toppari, J.; Meyts, E.R.D.; Leffers, H.; Juul, A.; Jensen, T.K.; Skakkebæk, N.E. Adverse trends in male reproductive health: We may have reached a crucial ‘tipping point’. Int. J. Androl. 2008, 31, 74–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, K.P.; Tanphaichitr, N. Human exposure to endocrine disrupters and semen quality. J. Toxicol. Environ. Health B Crit. Rev. 2008, 11, 188–220. [Google Scholar] [CrossRef]
- Lund, L.; Engebjerg, M.C.; Pedersen, L.; Ehrenstein, V.; Nørgaard, M.; Sørensen, H.F. Prevalence of hypospadias in Danish boys: A longitudinal study, 1977–2005. Eur. Urol. 2009, 55, 1022–1026. [Google Scholar] [CrossRef]
- Toppari, J.; Virtanen, H.E.; Main, K.M.; Skakkebaek, N.E. Cryptorchidism and hypospadias as a sign of testicular dysgenesis syndrome (TDS): Environmental connection. Birth Defects Res. A Clin. Mol. Teratol. 2010, 88, 910–919. [Google Scholar] [CrossRef] [PubMed]
- Mascarenhas, M.N.; Flaxman, S.R.; Boerma, T.; Vanderpoel, S.; Stevens, G.A. National, regional, and global trends in infertility prevalence since 1990: A systematic analysis of 277 health surveys. PLoS Med. 2012, 9, e1001356. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Mulgund, A.; Hamada, A.; Chyatte, M.R. A unique view on male infertility around the globe. Reprod. Biol. Endocrinol. 2015, 13, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inhorn, M.C.; Patrizio, P. Infertility around the globe: New thinking on gender, reproductive technologies and global movements in the 21st century. Hum. Reprod. Update 2015, 21, 411–426. [Google Scholar] [CrossRef] [Green Version]
- Skakkebaek, N.E.; Rajpert-De Meyts, E.; Buck Louis, G.M.; Toppari, J.; Andersson, A.M.; Eisenberg, M.L.; Jensen, T.K.; Jørgensen, N.; Swan, S.H.; Sapra, K.J.; et al. Male reproductive disorders and fertility trends: Influences of environment and genetic susceptibility. Physiol. Rev. 2016, 96, 55–97. [Google Scholar] [CrossRef] [Green Version]
- Landrigan, P.J.; Fuller, R.; Acosta, N.J.R.; Adeyi, O.; Arnold, R.; Basu, N.N.; Baldé, A.B.; Bertollini, R.; Bose-O’Reilly, S.; Boufford, J.I.; et al. Lancet Commission on pollution and health. Lancet 2018, 391, 462–512. [Google Scholar] [CrossRef] [Green Version]
- EFSA Scientific Committee. Scientific Opinion on the Hazard Assessment of Endocrine Disruptors: Scientific Criteria for Identification of Endocrine Disruptors and Appropriateness of Existing Test Methods for Assessing Effects Mediated by These Substances on Human Health and the Environment. EFSA J. 2013, 11, 3132. Available online: www.efsa.europa.eu/efsajournal (accessed on 10 April 2021). [CrossRef]
- Bergman, Å.; Heindel, J.J.; Jobling, S.; Kidd, K.A.; Zoeller, R.T. The State-of-the-Science of Endocrine Disrupting Chemicals; World Health Organization and United Nations Environment Programme Report–2012; WHO: Geneva, Switzerland, 2013; Available online: www.who.int/ceh/publications/endocrine/en/index.html (accessed on 10 April 2021).
- Kime, D.E. Endocrine Disruptors in Fish; Kluwer Academic Publishers: Boston, MA, USA, 1998. [Google Scholar]
- Kah, O. Endocrine targets of the hypothalamus and pituitary. In Fish Physiology: Fish Neuroendocrinology; Bernier, N.J., Van Der Kraak, G., Farrell, A.P., Brauner, C.J., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2009; Volume 28, pp. 75–112. [Google Scholar] [CrossRef]
- Schulz, R.W.; de França, L.R.; Lareyre, J.J.; Le Gac, F.; Chiarini-Garcia, H.; Nobrega, R.H.; Miura, T. Spermatogenesis in fish. Gen. Comp. Endocrinol. 2010, 165, 390–411. [Google Scholar] [CrossRef]
- Dufour, S.; Quérat, B.; Tostivint, H.; Pasqualini, C.; Vaudry, H.; Rousseau, K. Origin and evolution of the neuroendocrine control of reproduction in vertebrates, with special focus on genome and gene duplications. Physiol. Rev. 2020, 100, 869–943. [Google Scholar] [CrossRef]
- Gregory, M.; Aravindakshan, J.; Nadzialek, S.; Cyr, D.G. Effects of endocrine disrupting chemicals on testicular functions. In Fish Spermatology; Alavi, S.M.H., Cosson, J., Coward, K., Rafiee, G., Eds.; Alpha Science Ltd.: Oxford, UK, 2008; pp. 161–214. [Google Scholar]
- Ankley, G.T.; Bencic, D.C.; Breen, M.S.; Collette, T.W.; Conolly, R.B.; Denslow, N.D.; Edwards, S.W.; Ekman, D.R.; Garcia-Reyero, N.; Jensen, K.M.; et al. Endocrine disrupting chemicals in fish: Developing exposure indicators and predictive models of effects based on mechanism of action. Aquat. Toxicol. 2009, 92, 168–178. [Google Scholar] [CrossRef]
- Scholz, S.; Klüver, N. Effects of endocrine disrupters on sexual, gonadal development in fish. Sex. Dev. 2009, 3, 136–151. [Google Scholar] [CrossRef] [PubMed]
- Bosker, T.; Munkittrick, K.R.; MacLatchy, D.L. Challenges and opportunities with the use of biomarkers to predict reproductive impairment in fishes exposed to endocrine disrupting substances. Aquat. Toxicol. 2011, 100, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Leet, J.K.; Gall, H.E.; Sepúlveda, M.S. A review of studies on androgen and estrogen exposure in fish early life stages: Effects on gene and hormonal control of sexual differentiation. J. Appl. Toxicol. 2011, 31, 379–398. [Google Scholar] [CrossRef]
- Mennigen, J.A.; Stroud, P.; Zamora, J.M.; Moon, T.W.; Trudeau, V.L. Pharmaceuticals as neuroendocrine disruptors: Lessons learned from fish on prozac. J. Toxicol. Environ. Health. B Crit. Rev. 2011, 14, 387–412. [Google Scholar] [CrossRef]
- Hano, T. Studies on the evaluation of the effects of endocrine disrupting chemicals using transgenic see-through medaka (Oryzias latipes), olvas-GFP/STII-YI strain. Bull. Fish. Res. Agency 2012, 36, 1–56. [Google Scholar]
- Abdel-Moneim, A.; Coulter, D.P.; Mahapatra, C.T.; Sepúlveda, M.S. Intersex in fishes and amphibians: Population implications, prevalence, mechanisms and molecular biomarkers. J. Appl. Toxicol. 2015, 35, 1228–1240. [Google Scholar] [CrossRef]
- Golshan, M.; Alavi, S.M.H. Androgen signaling in male fishes: Examples of anti-androgenic chemicals that cause reproductive disorders. Theriogenology 2019, 139, 58–71. [Google Scholar] [CrossRef]
- Hatef, A.; Alavi, S.M.H.; Golshan, M.; Linhart, O. Toxicity of environmental contaminants to fish spermatozoa functions in vitro—A review. Aquat. Toxicol. 2013, 140–141, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Carnevali, O.; Santangeli, S.; Forner-Piquer, I.; Basili, D.; Maradonna, F. Endocrine disrupting chemicals in aquatic environment: What are the risks for fish gametes? Fish Physiol. Biochem. 2018, 44, 1561–1576. [Google Scholar] [CrossRef] [PubMed]
- Alavi, S.M.H.; Linhart, O.; Coward, K.; Rodina, M. Fish spermatology: Implication for aquaculture management. In Fish Spermatology; Alavi, S.M.H., Cosson, J., Coward, K., Rafiee, G., Eds.; Alpha Science Ltd.: Oxford, UK, 2008; pp. 397–460. [Google Scholar]
- Dzyuba, V.; Shelton, W.L.; Kholodnyy, V.; Boryshpolets, S.; Cosson, J.; Dzyuba, B. Fish sperm biology in relation to urogenital system structure. Theriogenology 2019, 132, 153–163. [Google Scholar] [CrossRef]
- Grier, H.J. Cellular organization of the testis and spermatogenesis in fishes. Am. Zool. 1981, 21, 345–357. [Google Scholar] [CrossRef] [Green Version]
- Billard, R. Spermatogenesis and spermatology of some teleost fish species. Reprod. Nutr. Dev. 1986, 2, 877–920. [Google Scholar] [CrossRef]
- Nagahama, Y. The functional morphology of teleosts gonads. In Fish Physiology, Reproduction Part A.; Hoar, W.S., Randall, D.J., Donaldson, E.M., Eds.; Academic Press Inc.: London, UK, 1983; Volume IX, pp. 223–275. [Google Scholar]
- Parenti, L.R.; Grier, H.J. Evolution and phylogeny of gonad morphology in bony fishes. Integr. Comp. Biol. 2004, 44, 333–348. [Google Scholar] [CrossRef] [Green Version]
- Lahnsteiner, F.; Patzner, R.A. The spermatic duct of blenniid fish (Teleostei, Blenniidae): Fine structure, histochemistry and function. Zoomorphology 1990, 110, 63–73. [Google Scholar] [CrossRef]
- Lahnsteiner, F.; Patzner, R.A.; Weismann, T. The spermatic ducts of salmonid fishes (Salmonidae, Teleostei). Morphology, histochemistry and composition of the secretion. J. Fish Biol. 1993, 42, 79–93. [Google Scholar] [CrossRef]
- Miura, T.; Yamauchi, K.; Takahashi, H.; Nagahama, Y. The role of hormone in the acquisition of sperm motility in salmonid fish. J. Exp. Zool. 1992, 261, 359–363. [Google Scholar] [CrossRef]
- Morisawa, S.; Ishida, K.; Okuno, M.; Morisawa, M. Roles of pH and cyclic adenosine monophosphate in the acquisition of potential for sperm motility during migration from the sea to the river in chum salmon. Mol. Reprod. Dev. 1993, 34, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Vizziano, D.; Fostier, A.; Loir, M.; Le Gac, F. Testis development, its hormonal regulation and spermiation induction in teleost fish. In Fish Spermatology; Alavi, S.M.H., Cosson, J., Coward, K., Rafiee, G., Eds.; Alpha Science Ltd.: Oxford, UK, 2008; pp. 103–140. [Google Scholar]
- Watanabe, A.; Onitake, K. The regulation of spermatogenesis in fish. In Fish Spermatology; Alavi, S.M.H., Cosson, J., Coward, K., Rafiee, G., Eds.; Alpha Science Ltd.: Oxford, UK, 2008; pp. 141–160. [Google Scholar]
- Jamieson, B.G.M. Fish Evolution and Systematics: Evidence from Spermatozoa; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Hara, M.; Okiyama, M. An ultrastructural review on the spermatozoa of Japanese fishes. Bull. Oce. Res. Inst. Univ. Tokyo 1998, 33, 1–138. [Google Scholar]
- Lahnsteiner, F.; Patzner, R.A. Sperm morphology and ultrastructure in fish. In Fish Spermatology; Alavi, S.M.H., Cosson, J., Coward, K., Rafiee, G., Eds.; Alpha Science Ltd.: Oxford, UK, 2008; pp. 1–61. [Google Scholar]
- Ingermann, R.L. Energy metabolism and respiration in fish spermatozoa. In Fish Spermatology; Alavi, S.M.H., Cosson, J., Coward, K., Rafiee, G., Eds.; Alpha Science Ltd.: Oxford, UK, 2008; pp. 241–266. [Google Scholar]
- Inaba, K.; Shiba, K. Microscopic analysis of sperm movement: Links to mechanisms and protein components. Microscopy 2018, 67, 144–155. [Google Scholar] [CrossRef]
- Morisawa, S.; Cherr, G.N. Acrosome reaction in spermatozoa from hagfish (Agnatha) Eptatretus burgeri and Eptatretus stouti: Acrosomal exocytosis and identification of filamentous actin. Dev. Growth Differ. 2002, 44, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Alavi, S.M.H.; Hatef, A.; Pšenička, M.; Kašpar, V.; Boryshpolets, S.; Dzyuba, B.; Cosson, J.; Bondarenko, V.; Rodina, M.; Gela, D.; et al. Sperm biology and control of reproduction in sturgeon: (II) Sperm morphology, acrosome reaction, motility and cryopreservation. Rev. Fish Biol. Fish. 2012, 22, 861–886. [Google Scholar] [CrossRef]
- Hatef, A.; Alavi, S.M.H.; Rodina, M.; Linhart, O. Morphology and fine structure of the Russian sturgeon, Acipenser gueldenstaedtii (Acipenseridae, Chondrostei) spermatozoa. J. Appl. Ichthyol. 2012, 28, 978–983. [Google Scholar] [CrossRef]
- Alavi, S.M.H.; Butts, I.A.E.; Hatef, A.; Mommens, M.; Trippel, E.A.; Litvak, M.K.; Babiak, I. Sperm morphology, ATP content and analysis of motility in Atlantic halibut (Hippoglossus hippoglossus L.). Can. J. Zool. 2011, 89, 219–228. [Google Scholar] [CrossRef]
- Inaba, K. Molecular architecture of sperm flagella: Molecules for motility and signaling. Zool. Sci. 2003, 20, 1043–1056. [Google Scholar] [CrossRef]
- Ciereszko, A. Chemical composition of seminal plasma and its physiological relationship with sperm motility, fertilizing capacity and cryopreservation success in fish. In Fish Spermatology; Alavi, S.M.H., Cosson, J., Coward, K., Rafiee, G., Eds.; Alpha Science Ltd.: Oxford, UK, 2008; pp. 215–240. [Google Scholar]
- Kowalski, R.K.; Cejko, B.I. Sperm quality in fish: Determinants and affecting factors. Theriogenology 2019, 135, 94–108. [Google Scholar] [CrossRef]
- Morisawa, M.; Suzuki, K. Osmolality and potassium ions: Their roles in initiation of sperm motility in teleosts. Science 1980, 210, 1145–1147. [Google Scholar] [CrossRef]
- Morisawa, M.; Suzuki, K.; Morisawa, S. Effects of potassium and osmolality on spermatozoan motility of salmonid fishes. J. Exp. Biol. 1983, 107, 105–113. [Google Scholar] [CrossRef]
- Billard, R. Reproduction in rainbow trout: Sex differentiation, dynamics of gametogenesis, biology and preservation of gametes. Aquaculture 1992, 100, 263–298. [Google Scholar] [CrossRef]
- Alavi, S.M.H.; Cosson, J.; Karami, M.; Amiri, B.M.; Akhoundzadeh, M.A. Spermatozoa motility in the Persian sturgeon, Acipenser persicus: Effects of pH, dilution rate, ions and osmolality. Reproduction 2004, 128, 819–828. [Google Scholar] [CrossRef]
- Morisawa, M.; Oda, S.; Yoshida, M.; Takai, H. Transmembrane signal transduction for the regulation of sperm motility in fishes and ascidians. In The Male Gamete: From Basic to Clinical Applications; Gagnon, C., Ed.; Cache River Press: Vienna, IL, USA, 1999; pp. 149–160. [Google Scholar]
- Alavi, S.M.H.; Cosson, J. Sperm motility in fishes: (II) Effects of ions and osmotic pressure. Cell Biol. Int. 2006, 30, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Morisawa, M. Adaptation and strategy for fertilization in the sperm of teleosts fish. J. Appl. Ichthyol. 2008, 24, 362–370. [Google Scholar] [CrossRef]
- Alavi, S.M.H.; Cosson, J.; Bondarenko, O.; Linhart, O. Sperm motility in fishes: (III) diversity of regulatory signals from membrane to the axoneme. Theriogenology 2019, 136, 143–165. [Google Scholar] [CrossRef]
- Morisawa, M.; Okuno, M. Cyclic AMP induces maturation of trout sperm axoneme to initiate motility. Nature 1982, 295, 703–704. [Google Scholar] [CrossRef]
- Cosson, M.P.; Cosson, J.; Andre, F.; Billard, R. cAMP/ATPrelationshipinthe activation of trout sperm motility: Their interaction in membrane-deprived models and in live spermatozoa. Cell Motil. Cytoskelet. 1995, 31, 159–176. [Google Scholar] [CrossRef] [PubMed]
- Linhart, O.; Cosson, J.; Mims, S.D.; Shelton, W.L.; Rodina, M. Effects of ions on the motility of fresh and demembranated paddlefish (Polyodon spathula) spermatozoa. Reproduction 2002, 124, 713–719. [Google Scholar] [CrossRef]
- Tubbs, C.; Thomas, P. Progestin signaling through an olfactory G protein and membrane progestin receptor-a in Atlantic croaker sperm: Potential role in induction of sperm hypermotility. Endocrinology 2009, 150, 473–484. [Google Scholar] [CrossRef] [Green Version]
- Tan, W.; Aizen, J.; Thomas, P. Membrane progestin receptor-alpha mediates progestin-induced sperm hypermotility and increased fertilization success in southern flounder (Paralichthys lethostigma). Gen. Comp. Endocrinol. 2014, 200, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Alavi, S.M.H.; Cosson, J. Sperm motility in fishes: (I) Effects of pH and temperature. Cell Biol. Int. 2005, 29, 101–110. [Google Scholar] [CrossRef]
- Cosson, J. Frenetic activation of fish spermatozoa flagella entails short-term motility, portending their precocious decadence. J. Fish Biol. 2010, 76, 240–279. [Google Scholar] [CrossRef]
- Ziętara, M.S.; Biegniewska, A.; Rurangwa, E.; Swierczynski, J.; Ollevier, F.; Skorkowski, E.F. Bioenergetics of fish spermatozoa during semen storage. Fish Physiol. Biochem. 2009, 35, 607–614. [Google Scholar] [CrossRef]
- Dzyuba, B.; Bondarenko, O.; Fedorov, P.; Gazo, I.; Prokopchuk, G.; Cosson, J. Energetics of fish spermatozoa: The proven and the possible. Aquaculture 2017, 472, 60–72. [Google Scholar] [CrossRef]
- Lahnsteiner, F.; Patzner, R.A.; Weismann, T. Energy resources of spermatozoa of the rainbow trout Oncorhynchus mykiss (Pisces, Teleostei). Reprod. Nut. Dev. 1993, 33, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Saudrais, C.; Fierville, F.; Loir, M.; Le Remeur, E.; Cibert, C.; Cosson, J. The use of phosphocreatine plus ADP as energy source for motility of membrane-derived trout spermatozoa. Cell Motil. Cytoskel. 1998, 41, 91–106. [Google Scholar] [CrossRef]
- Woolsey, J.; Ingermann, R.L. Acquisition of the potential for sperm motility in steelhead (Oncorhynchus mykiss): Effect of pH on dynein ATPase. Fish Physiol. Biochem. 2003, 29, 47–56. [Google Scholar] [CrossRef]
- Kinsey, W.H.; Sharma, D.; Kinsey, S.C. Fertilization and egg activation in fishes. In The Fish Oocyte: From Basic Studies to Biotechnological Applications; Babin, P.J., Cerdà, J., Lubzens, E., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 397–409. [Google Scholar] [CrossRef]
- Kudo, S. Fertilization, cortical reaction, polyspermy preventing and anti-microbial mechanisms in fish eggs. Bull. Inst. Zool. Acad. Sinica 1991, 16, 313–340. [Google Scholar]
- Ginzburg, A.S. Fertilization in Fishes and Problem of Polyspermy; Israel Program for Scientific Translations; National Technical Information Service US Department of Commerce: Springfield, VA, USA, 1972.
- Iwamatsu, T. Stages of normal development in the medaka Oryzias latipes. Mech. Dev. 2004, 121, 605–618. [Google Scholar] [CrossRef]
- Tvedt, H.B.; Benfey, T.J.; Martin-Robichaud, D.J.; Power, J. The relationship between sperm density, spermatocrit, sperm motility and fertilization success in Atlantic halibut, Hippoglossus hippoglossus. Aquaculture 2001, 194, 191–200. [Google Scholar] [CrossRef]
- Rideout, R.M.; Trippel, E.A.; Litvak, M.K. Relationship between sperm density, spermatocrit, sperm motility and spawning date in wild and cultured haddock. J. Fish Biol. 2004, 65, 319–332. [Google Scholar] [CrossRef]
- Kaspar, V.; Kohlmann, K.; Vandeputte, M.; Rodina, M.; Gela, D.; Kocour, M.; Alavi, S.M.H.; Hulak, M.; Linhart, O. Equalizing sperm concentrations in a common carp (Cyprinus carpio) sperm pool does not affect variance in proportions of larvae sired in competition. Aquaculture 2007, 272 (Suppl. S1), S204–S209. [Google Scholar] [CrossRef]
- Hatef, A.; Niksirat, H.; Alavi, S.M.H. Composition of ovarian fluid in endangered Caspian brown trout, Salmo trutta caspius, and its effects on spermatozoa motility and fertilizing ability compared to freshwater and a saline medium. Fish Physiol. Biochem. 2009, 35, 695–700. [Google Scholar] [CrossRef]
- Butts, I.A.E.; Trippel, E.A.; Litvak, M.K. The effect of sperm to egg ratio and gamete contact time on fertilization success in Atlantic cod Gadus morhua. Aquaculture 2009, 286, 89–94. [Google Scholar] [CrossRef]
- Butts, I.A.E.; Roustaian, P.; Litvak, M.K. Fertilization strategies for winter flounder: Effects of spermatozoa density and the duration of gamete receptivity. Aquac. Biol. 2012, 16, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Linhart, O.; Cheng, Y.; Xin, M.M.; Rodina, M.; Tučková, V.; Shelton, W.L.; Kašpar, V. Standardization of egg activation and fertilization in sterlet (Acipenser ruthenus). Aquac. Rep. 2020, 17, 100381. [Google Scholar] [CrossRef]
- Cheng, Y.; Franek, R.; Rodina, M.; Xin, M.; Cosson, J.; Zhang, S.; Linhart, O. Optimization of Sperm Management and Fertilization in Zebrafish (Danio rerio (Hamilton)). Animals 2021, 11, 1558. [Google Scholar] [CrossRef] [PubMed]
- Linhart, O.; Rodina, M.; Flajšhans, M.; Mavrodiev, N.; Nebesárová, J.; Gela, D.; Kocour, M. Studies on sperm of diploid and triploid tench, Tinca tinca (L.). Aquac. Int. 2006, 14, 9–25. [Google Scholar] [CrossRef]
- Piferrer, F.; Beaumont, A.; Falguière, J.C.; Flajšhans, M.; Haffray, P.; Colombo, L. Polyploid fish and shellfish: Production, biology and applications to aquaculture for performance improvement and genetic containment. Aquaculture 2009, 293, 125–156. [Google Scholar] [CrossRef] [Green Version]
- Alavi, S.M.H.; Drozd, B.; Hatef, A.; Flajšhans, M. Sperm morphology, motility and velocity in naturally polyploid European weatherfish (Misgurnus fossilis L.). Theriogenology 2013, 80, 153–160. [Google Scholar] [CrossRef]
- Pšenička, M.; Flajšhans, M.; Hulák, M.; Kašpar, V.; Rodina, M.; Borishpolets, S.; Linhart, O. The influence of ploidy level on ultrastructure and motility of tench Tinca tinca (L.) spermatozoa. Rev. Fish Biol. Fish. 2010, 20, 331–338. [Google Scholar] [CrossRef]
- Pšenička, M.; Kašpar, V.; Rodina, M.; Gela, D.; Hulák, M.; Flajšhans, M. Comparative study on ultrastructure and motility parameters of spermatozoa of tetraploid and hexaploid Siberian sturgeon Acipenser baerii. J. Appl. Ichthyol. 2011, 27, 683–686. [Google Scholar] [CrossRef]
- Gage, M.J.G.; MacFarlane, C.; Yeates, S.; Shackleton, R.; Parker, G.A. Relationships between sperm morphometry and sperm motility in the Atlantic salmon. J. Fish Biol. 2002, 60, 1528–1539. [Google Scholar] [CrossRef]
- Alavi, S.M.H.; Psenicka, M.; Rodina, M.; Policar, T.; Linhart, O. Changes of sperm morphology, volume, density and motility and seminal plasma composition in Barbus barbus (Cyprinidae: Teleostei) during the reproductive season. Aquat. Living Resour. 2008, 21, 75–80. [Google Scholar] [CrossRef]
- Alavi, S.M.H.; Rodina, M.; Viveiros, A.T.M.; Cosson, J.; Gela, D.; Boryshpolets, S.; Linhart, O. Effects of osmolality on sperm morphology, motility and flagellar wave parameters in Northern pike (Esox lucius L.). Theriogenology 2009, 72, 32–43. [Google Scholar] [CrossRef]
- Ishijima, S.; Hara, M.; Okiyama, M. Comparative studies on spermatozoan motility of Japanese fishes. Bull. Oce. Res. Inst. Univ. Tokyo 1998, 33, 139–152. [Google Scholar]
- Beirão, J.; Boulais, M.; Gallego, V.; O’Brien, J.K.; Peixoto, S.; Robeck, T.R.; Cabrita, E. Sperm handling in aquatic animals for artificial reproduction. Theriogenology 2019, 133, 161–178. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, E.; Lee-Estévez, M.; Valdebenito, I.; Farías, J.G.; Romero, J. Potential biomarkers of DNA quality in cryopreserved fish sperm: Impact on gene expression and embryonic development. Rev. Aquac. 2020, 12, 382–391. [Google Scholar] [CrossRef]
- Kim, D.S.; Jo, J.Y.; Lee, T.Y. Induction of triploidy in mud loach (Misgurnus mizolepis) and its effect on gonad development and growth. Aquaculture 1994, 120, 263–270. [Google Scholar] [CrossRef]
- Chourrout, D.; Chevassus, B.; Krieg, F.; Happe, A.; Burger, G.; Renard, P. Production of second generation triploid and tetraploid rainbow trout by mating tetraploid males and diploid females-potential of tetraploid fish. Theor. Appl. Genet. 1986, 72, 193–206. [Google Scholar] [CrossRef]
- Oshima, K.; Morishima, K.; Yamaha, E.; Arai, K. Reproductive capacity of triploid loaches obtained from Hokkaido Island, Japan. Ichthyol. Res. 2005, 52, 1–8. [Google Scholar] [CrossRef]
- Fujimoto, T.; Yasui, G.S.; Yoshikawa, H.; Yamaha, E.; Arai, K. Genetic and reproductive potential of spermatozoa of diploid and triploid males obtained from interspecific hybridization of Misgurnus anguillicaudatus female with M. mizolepis male. J. Appl. Ichthyol. 2008, 24, 430–437. [Google Scholar] [CrossRef] [Green Version]
- Kawamura, K.; Ueda, T.; Aoki, K.; Hosoya, K. Spermatozoa in triploids of the rosy bitterling, Rhodeus ocellatus ocellatus. J. Fish Biol. 1999, 55, 420–432. [Google Scholar] [CrossRef]
- Cabrita, E.; Robles, V.; Rebordinos, L.; Sarasquete, C.; Herraez, M.P. Evaluation of DNA damage in rainbow trout (Oncorhynchus mykiss) and gilthead sea bream (Sparus aurata) cryopreserved sperm. Cryobiology 2005, 50, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Shaliutina, A.; Hulak, M.; Gazo, I.; Linhartova, P.; Linhart, O. Effect of short-term storage on quality parameters, DNA integrity, and oxidative stress in Russian (Acipenser gueldenstaedtii) and Siberian (Acipenser baerii) sturgeon sperm. Anim. Reprod. Sci. 2013, 139, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Linhart, O.; Rodina, M.; Gela, D.; Kocour, M.; Vandeputte, M. Spermatozoal competition in common carp (Cyprinus carpio): What is the primary determinant of competition success? Reproduction 2005, 130, 705–711. [Google Scholar] [CrossRef] [Green Version]
- Gage, M.J.G.; MacFarlane, C.P.; Yeates, S.; Ward, R.G.; Searle, J.B.; Parker, G.A. Spermatozoal traits and sperm competition in Atlantic salmon: Relative sperm velocity is the primary determinant of fertilization success. Curr. Biol. 2004, 14, 44–47. [Google Scholar] [CrossRef]
- Linhart, O.; Alavi, S.M.H.; Rodina, M.; Gela, D.; Cosson, J. Comparison of sperm velocity, motility and fertilizing ability between firstly and secondly activated spermatozoa of common carp (Cyprinus carpio). J. Appl. Ichthyol. 2008, 24, 386–392. [Google Scholar] [CrossRef]
- Wilson-Leedy, J.G.; Ingermann, R.L. Development of a novel CASA system based on open source software for characterization of zebrafish sperm motility parameters. Theriogenology 2007, 67, 661–672. [Google Scholar] [CrossRef]
- Cosson, J. Methods to analyse the movements of the spermatozoa and their flagella. In Fish Spermatology; Alavi, S.M.H., Cosson, J., Coward, K., Rafiee, G., Eds.; Alpha Science Ltd.: Oxford, UK, 2008; pp. 63–102. [Google Scholar]
- Amann, R.P.; Waberski, D. Computer-assisted sperm analysis (CASA): Capabilities and potential developments. Theriogenology 2014, 81, 5–17. [Google Scholar] [CrossRef]
- McMaster, M.E.; Van Der Kraak, G.J.; Portt, C.B.; Munkittrick, K.R.; Sibley, P.K.; Smith, I.R.; Dixon, D.G. Changes in hepatic mixed function oxygenase (MFO) activity, plasma steroid levels, and age at maturity of a white sucker (Catostomus commersoni) population exposed to bleached kraft pulp mill effluent. Aquat. Toxicol. 1991, 21, 199–217. [Google Scholar] [CrossRef]
- Munkittrick, K.R.; Portt, C.B.; Van Der Kraak, G.J.; Smith, I.R.; Rokosh, D. Impact of bleached kraft mill effluent on population characteristics, liver MFO activity and serum steroid levels of a Lake Superior white sucker (Catostomus commersoni) population. Can. J. Fish. Aquat. Sci. 1991, 48, 1371–1380. [Google Scholar] [CrossRef]
- Munkittrick, K.R.; McMaster, M.E.; Portt, C.B.; Van Der Kraak, G.J.; Smith, I.R.; Dixon, D.G. Changes in maturity, plasma sex steroid levels, hepatic mixed-function oxygenase activity and the presence of external lesions in lake whitefish (Coregonus clupeaformis) exposed to bleached kraft mill effluent. Can. J. Fish. Aquat. Sci. 1992, 49, 1560–1569. [Google Scholar] [CrossRef]
- Allen, Y.; Scott, A.P.; Matthiessen, P.; Haworth, S.; Thain, J.E.; Feist, S. Survey of estrogenic activity in United Kingdom estuarine and coastal waters and its effects on gonadal development of the flounder Platichthys flesus. Environ. Toxicol. Chem. 1999, 18, 1791–1800. [Google Scholar] [CrossRef]
- Harshbarger, J.C.; Coffey, M.J.; Young, M.Y. Inter-sexes in Mississippi River shovelnose sturgeon sampled below Saint Louis, Missouri, USA. Mar. Environ. Res. 2000, 50, 247–250. [Google Scholar] [CrossRef]
- Hashimoto, S.; Bessho, H.; Hara, A.; Nakamura, M.; Iguchi, T.; Fujita, K. Elevated serum vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) from Tokyo Bay, Japan. Mar. Environ. Res. 2000, 49, 37–53. [Google Scholar] [CrossRef]
- Viganò, L.; Arillo, A.; Bottero, S.; Massari, A.; Mandich, A. First observation of intersex cyprinids in the Po River (Italy). Sci. Total Environ. 2001, 269, 189–194. [Google Scholar] [CrossRef]
- Jobling, S.; Beresford, N.; Nolan, M.; Rodgers-Gray, T.; Brighty, G.C.; Sumpter, J.P.; Tyler, C.R. Altered sexual maturation and gamete production in wild roach (Rutilus rutilus) living in rivers that receive treated sewage effluents. Biol. Reprod. 2002, 66, 272–281. [Google Scholar] [CrossRef]
- Sepúlveda, M.S.; Johnson, W.E.; Higman, J.C.; Denslow, N.D.; Schoeb, T.R.; Gross, T.S. An evaluation of biomarkers of reproductive function and potential contaminant effects in Florida largemouth bass (Micropterus salmoides floridanus) sampled from the St. Johns River. Sci. Total Environ. 2002, 289, 133–144. [Google Scholar] [CrossRef]
- Aravindakshan, J.; Paquet, V.; Gregory, M.; Dufresne, J.; Fournier, M.; Marcogliese, D.J.; Cyr, D.J. Consequences of xenoestrogen exposure on male reproductive function in Spottail Shiners (Notropis hudsonius). Toxicol. Sci. 2004, 78, 156–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavanagh, R.J.; Balch, G.C.; Kiparissis, Y.; Niimi, A.J.; Sherry, J.; Tinson, C.; Metcalfe, C.D. Endocrine disruption and altered gonadal development in white perch (Morone americana) from the lower Great Lakes region. Environ. Health Perspect. 2004, 112, 898–902. [Google Scholar] [CrossRef] [Green Version]
- Bjerregaard, L.B.; Korsgaard, B.; Bjerregaard, P. Intersex in wild roach (Rutilus rutilus) from Danish sewage effluent-receiving streams. Ecotoxicol. Environ. Saf. 2006, 64, 321–328. [Google Scholar] [CrossRef]
- Woodling, J.D.; Lopez, E.M.; Maldonado, T.A.; Norris, D.O.; Vajda, A.M. Intersex and other reproductive disruption of fish in wastewater effluent dominated Colorado streams. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 144, 10–15. [Google Scholar] [CrossRef]
- Marchand, M.J.; Pieterse, G.M.; Barnhoorn, I.E.J. Preliminary results on sperm motility and testicular histology of two feral fish species, Oreochromis mossambicus and Clarias gariepinus, from a currently DDT-sprayed area, South Africa. J. Appl. Ichthyol. 2008, 24, 423–429. [Google Scholar] [CrossRef]
- Marchand, M.J.; Pieterse, G.M.; Barnhoorn, I.E.J. Sperm motility and testicular histology as reproductive indicators of fish health of two feral fish species from a currently DDT sprayed area, South Africa. J. Appl. Ichthyol. 2010, 26, 707–714. [Google Scholar] [CrossRef]
- Jeffries, K.M.; Nelson, E.R.; Jackson, L.J.; Habibi, H.R. Basin-wide impacts of compounds with estrogen-like activity on longnose dace (Rhinichthys cataractae) in two Prairie Rivers of Alberta, Canada. Environ. Toxicol. Chem. 2008, 27, 2042–2052. [Google Scholar] [CrossRef]
- Jeffries, K.M.; Jackson, L.J.; Ikonomou, M.G.; Habibi, H.R. Presence of natural and anthropogenic organic contaminants and potential fish health impacts along two river gradients in Alberta, Canada. Environ. Toxicol. Chem. 2010, 29, 2379–2387. [Google Scholar] [CrossRef] [PubMed]
- Barnhoorn, I.E.J.; van Dyk, J.C.; Pieterse, G.M.; Bornman, M.S. Intersex in feral indigenous freshwater Oreochromis mossambicus, from various parts in the Luvuvhu River, Limpopo Province, South Africa. Ecotoxicol. Environ. Saf. 2010, 73, 1537–1542. [Google Scholar] [CrossRef] [PubMed]
- Solé, M.; Raldúa, D.; Piferrer, F.; Barceló, D.; Porte, C. Feminization of wild carp, Cyprinus carpio, in a polluted environment: Plasma steroid hormones, gonadal morphology and xenobiotic metabolizing system. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2003, 136, 145–156. [Google Scholar] [CrossRef]
- Vajda, A.M.; Barber, L.B.; Gray, J.L.; Lopez, E.M.; Woodling, J.D.; Norris, D.O. Reproductive disruption in fish downstream from an estrogenic wastewater effluent. Environ. Sci. Technol. 2008, 42, 3407–3414. [Google Scholar] [CrossRef]
- Gilroy, E.A.M.; McMaster, M.E.; Parrott, J.L.; Hewitt, L.M.; Park, B.J.; Brown, S.B.; Sherry, J.P. Assessment of the health status of wild fish from the Wheatley Harbour Area of Concern, Ontario, Canada. Environ. Toxicol. Chem. 2012, 31, 2798–2811. [Google Scholar] [CrossRef]
- Bahamonde, P.A.; Tetreault, G.R.; McMaster, M.E.; Servos, M.R.; Martyniuk, C.J.; Munkittrick, K.R. Molecular signatures in rainbow darter (Etheostoma caeruleum) inhabiting an urbanized river reach receiving wastewater effluents. Aquat. Toxicol. 2014, 148, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Bahamonde, P.A.; Fuzzen, M.L.; Bennett, C.J.; Tetreault, G.R.; McMaster, M.E.; Servos, M.R.; Martyniuk, C.J.; Munkittrick, K.R. Whole organism responses and intersex severity in rainbow darter (Etheostoma caeruleum) following exposures to municipal wastewater in the Grand River basin, ON, Canada. Part A. Aquat. Toxicol. 2015, 159, 290–301. [Google Scholar] [CrossRef] [PubMed]
- McMaster, M.E.; Portt, C.B.; Munkittrick, K.R.; Dixon, D.G. Milt characteristics, reproductive performance, and larval survival and development of white sucker exposed to bleached kraft mill effluent. Ecotoxicol. Environ. Saf. 1992, 23, 103–117. [Google Scholar] [CrossRef]
- Jobling, S.; Coey, S.; Whitmore, J.G.; Kime, D.E.; Van Look, K.J.W.; McAllister, B.G.; Beresford, N.; Henshaw, A.C.; Brighty, G.; Tyler, C.R.; et al. Wild intersex roach (Rutilus rutilus) have reduced fertility. Biol. Reprod. 2002, 67, 515–524. [Google Scholar] [CrossRef]
- Wagenaar, G.; Botha, T.; Barnhoorn, I. Sperm motility and testicular histology as reproductive indicators in Clarias gariepinus from an eutrophic impoundment, South Africa. J. Appl. Ichthyol. 2012, 28, 990–997. [Google Scholar] [CrossRef]
- Fuzzen, M.L.M.; Bennett, C.J.; Tetreault, G.R.; McMaster, M.E.; Servos, M.R. Severe intersex is predictive of poor fertilization success in populations of rainbow darter (Etheostoma caeruleum). Aquat. Toxicol. 2015, 160, 106–116. [Google Scholar] [CrossRef]
- Jenkins, J.A.; Olivier, H.M.; Draugelis-Dale, R.O.; Eilts, B.E.; Torres, L.; Patiño, R.; Nilsen, E.; Goodbred, S.L. Assessing reproductive and endocrine parameters in male largescale suckers (Catostomus macrocheilus) along a contaminant gradient in the lower Columbia River, USA. Sci. Total Environ. 2014, 484, 365–378. [Google Scholar] [CrossRef] [Green Version]
- Agbohessi, P.T.; Toko, I.I.; Ouédraogo, A.; Jauniaux, T.; Mandiki, S.N.M.; Kestemont, P. Assessment of the health status of wild fish inhabiting a cotton basin heavily impacted by pesticides in Benin (West Africa). Sci. Total Environ. 2015, 506-507, 567–584. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, J.A.; Rosen, M.R.; Draugelis-Dale, R.O.; Echols, K.R.; Torres, L.; Wieser, C.M.; Kersten, C.A.; Goodbred, S.L. Sperm quality biomarkers complement reproductive and endocrine parameters in investigating environmental contaminants in common carp (Cyprinus carpio) from the Lake Mead National Recreation Area. Environ. Res. 2018, 163, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Van der Kraak, G.J.; Munkittrick, K.R.; McMaster, M.E.; Portt, C.B.; Chang, J.P. Exposure to bleached kraft pulp mill effluent disrupts the pituitary-gonadal axis of white sucker at multiple sites. Toxicol. Appl. Pharmacol. 1992, 115, 224–233. [Google Scholar] [CrossRef]
- Flammarion, P.; Brion, F.; Babut, M.; Garric, J.; Migeon, B.; Noury, P.; Thybaud, E.; Palazzi, X.; Tyler, C.R. Induction of fish vitellogenin and alterations in testicular structure: Preliminary results of estrogenic effects in Chub (Leuciscus cephalus). Ecotoxicology 2000, 9, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Randák, T.; Žlábek, V.; Pulkrabová, J.; Kolářová, J.; Kroupová, H.; Široká, Z.; Velíšek, J.; Svobodová, Z.; Hajšlová, J. Effects of pollution on chub in the River Elbe, Czech Republic. Ecotoxicol. Environ. Saf. 2009, 72, 737–746. [Google Scholar] [CrossRef]
- Celander, M.C. Cocktail effects on biomarker responses in fish. Aquat. Toxicol. 2011, 105, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Hamid, N.; Junaid, M.; Pei, D.S. Combined toxicity of endocrine-disrupting chemicals: A review. Ecotoxicol. Environ. Saf. 2021, 215, 112136. [Google Scholar] [CrossRef]
- Pawlowski, S.; Sauer, A.; Shears, J.A.; Tyler, C.R.; Braunbeck, T. Androgenic and estrogenic effects of the synthetic androgen 17α-methyltestosterone on sexual development and reproductive performance in the fathead minnow (Pimephales promelas) determined using the gonadal recrudescence assay. Aquat. Toxicol. 2004, 68, 277–291. [Google Scholar] [CrossRef]
- Pawlowski, S.; van Aerle, R.; Tyler, C.R.; Braunbeck, T. Effects of 17α-ethinylestradiol in a fathead minnow (Pimephales promelas) gonadal recrudescence assay. Ecotoxicol. Environ. Saf. 2004, 57, 330–345. [Google Scholar] [CrossRef]
- Kang, I.J.; Yokota, H.; Oshima, Y.; Tsuruda, Y.; Shimasaki, Y.; Honjo, T. The effects of methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias latipes). Aquat. Toxicol. 2008, 87, 37–46. [Google Scholar] [CrossRef]
- Lahnsteiner, F.; Berger, B.; Grubinger, F.; Weismann, T. The effect of 4-nonylphenol on semen quality, viability of gametes, fertilization success, and embryo and larvae survival in rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 2005, 71, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Kang, I.J.; Yokota, H.; Oshima, Y.; Tsuruda, Y.; Oe, T.; Imada, N.; Tadokoro, H.; Honjo, T. Effects of bisphenol a on the reproduction of Japanese medaka (Oryzias latipes). Environ. Toxicol. Chem. 2002, 21, 2394–2400. [Google Scholar] [CrossRef]
- Martinović, D.; Blake, L.S.; Durhan, E.J.; Greene, K.J.; Kahl, M.D.; Jensen, K.M.; Makynen, E.A.; Villeneuve, D.L.; Ankley, G.T. Reproductive toxicity of vinclozolin in the fathead minnow: Confirming an anti-androgenic mode of action. Environ. Toxicol. Chem. 2008, 27, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Seki, M.; Yokota, H.; Matsubara, H.; Tsuruda, Y.; Maeda, M.; Tadokoro, H.; Kobayashi, K. Effect of ethinylestradiol on the reproduction and induction of vitellogenin and testis-ova in medaka (Oryzias latipes). Environ. Toxicol. Chem. 2002, 21, 1692–1698. [Google Scholar] [CrossRef]
- Hashimoto, S.; Watanabe, E.; Ikeda, M.; Terao, Y.; Strüssmann, C.A.; Inoue, M.; Hara, A. Effects of ethinylestradiol on medaka (Oryzias latipes) as measured by sperm motility and fertilization success. Arch. Environ. Contam. Toxicol. 2009, 56, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Devaux, A.; Fiat, L.; Gillet, C.; Bony, S. Reproduction impairment following paternal genotoxin exposure in brown trout (Salmo trutta) and Arctic charr (Salvelinus alpinus). Aquat. Toxicol. 2011, 101, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Kang, I.J.; Yokota, H.; Oshima, Y.; Tsuruda, Y.; Hano, T.; Maeda, M.; Imada, N.; Tadokoro, H.; Honjo, T. Effects of 4-nonylphenol on reproduction of Japanese medaka, Oryzias latipes. Environ. Toxicol. Chem. 2003, 22, 2438–2445. [Google Scholar] [CrossRef] [PubMed]
- Uren-Webster, T.M.; Lewis, C.; Filby, A.L.; Paull, G.C.; Santos, E.M. Mechanisms of toxicity of di (2-ethylhexyl) phthalate on the reproductive health of male zebrafish. Aquat. Toxicol. 2010, 99, 360–369. [Google Scholar] [CrossRef]
- Kang, I.J.; Yokota, H.; Oshima, Y.; Tsuruda, Y.; Yamaguchi, T.; Maeda, M.; Imada, N.; Tadokoro, H.; Honjo, T. Effect of 17β-estradiol on the reproduction of Japanese medaka (Oryzias latipes). Chemosphere 2002, 47, 71–80. [Google Scholar] [CrossRef]
- Schultz, I.R.; Skillman, A.; Nicolas, J.M.; Cyr, D.G.; Nagler, J.J. Short-term exposure to 17α-ethynylestradiol decreases the fertility of sexually maturing male rainbow trout (Oncorhynchus mykiss). Environ. Toxicol. Chem. 2003, 22, 1272–1280. [Google Scholar] [CrossRef]
- Lahnsteiner, F.; Berger, B.; Kletzl, M.; Weismann, T. Effect of 17β-estradiol on gamete quality and maturation in two salmonid species. Aquat. Toxicol. 2006, 79, 124–131. [Google Scholar] [CrossRef]
- Lahnsteiner, F.; Berger, B.; Kletzl, M.; Weismann, T. Effect of bisphenol A on maturation and quality of semen and eggs in the brown trout, Salmo trutta f. fario. Aquat.Toxicol. 2005, 75, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Li, Y.; Wang, W.; Wu, P.; Ru, S. Exposure to monocrotophos pesticide during sexual development causes the feminization/demasculinization of the reproductive traits and a reduction in the reproductive success of male guppies (Poecilia reticulata). Toxicol. Appl. Pharmacol. 2012, 263, 163–170. [Google Scholar] [CrossRef]
- Ji, K.; Hong, S.; Kho, Y.; Choi, K. Effects of bisphenol S exposure on endocrine functions and reproduction of zebrafish. Environ. Sci. Technol. 2013, 47, 8793–8800. [Google Scholar] [CrossRef]
- Shi, J.; Jiao, Z.; Zheng, S.; Li, M.; Zhang, J.; Feng, Y.; Yin, J.; Shao, B. Long-term effects of bisphenol AF (BPAF) on hormonal balance and genes of hypothalamus-pituitary-gonad axis and liver of zebrafish (Danio rerio), and the impact on offspring. Chemosphere 2015, 128, 252–257. [Google Scholar] [CrossRef]
- Montgomery, T.M.; Brown, A.C.; Gendelman, H.K.; Ota, M.; Clotfelter, E.D. Exposure to 17α-ethinylestradiol decreases motility and ATP in sperm of male fighting fish Betta splendens. Environ. Toxicol. 2014, 29, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Naderi, M.; Wong, M.Y.L.; Gholami, F. Developmental exposure of zebrafish (Danio rerio) to bisphenol-S impairs subsequent reproduction potential and hormonal balance in adults. Aquat. Toxicol. 2014, 148, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xiao, Y.; Gai, Z.; Li, R.; Zhu, Z.; Bai, C.; Tanguay, R.L.; Xu, X.; Huang, C.; Dong, Q. Reproductive toxicity of low level bisphenol A exposures in a two-generation zebrafish assay: Evidence of male-specific effects. Aquat. Toxicol. 2015, 169, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Saili, K.S.; Liu, Y.; Li, L.; Zhao, Y.; Jia, Y.; Bai, C.; Tanguay, R.L.; Dong, Q.; Huang, C. Developmental bisphenol A exposure impairs sperm function and reproduction in zebrafish. Chemosphere 2017, 169, 262–270. [Google Scholar] [CrossRef]
- Zhu, Y.; Hua, R.; Zhou, Y.; Li, H.; Quan, S.; Yu, Y. Chronic exposure to mono-(2-ethylhexyl)-phthalate causes endocrine disruption and reproductive dysfunction in zebrafish. Environ. Toxicol. Chem. 2016, 35, 2117–2124. [Google Scholar] [CrossRef]
- Corradetti, B.; Stronati, A.; Tosti, L.; Manicardi, G.; Carnevali, O.; Bizzaro, D. Bis-(2-ethylexhyl) phthalate impairs spermatogenesis in zebrafish (Danio rerio). Reprod. Biol. 2013, 13, 195–202. [Google Scholar] [CrossRef]
- Watanabe, H.; Horie, Y.; Takanobu, H.; Koshio, M.; Flynn, K.; Iguchi, T.; Tatarazako, N. Medaka extended one-generation reproduction test evaluating 4-nonylphenol. Environ. Toxicol. Chem. 2017, 36, 3254–3266. [Google Scholar] [CrossRef]
- Zhang, Y.; Guan, Y.; Zhang, T.; Yuan, C.; Liu, Y.; Wang, Z. Adult exposure to bisphenol A in rare minnow Gobiocypris rarus reduces sperm quality with disruption of testicular aquaporins. Chemosphere 2018, 193, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Kang, I.J.; Hano, T.; Oshima, Y.; Yokota, H.; Tsuruda, Y.; Shimasaki, Y.; Honjo, T. Anti-androgen flutamide affects gonadal development and reproduction in medaka (Oryzias latipes). Mar. Environ. Res. 2006, 62, S253–S257. [Google Scholar] [CrossRef]
- Schoenfuss, H.L.; Levitt, J.T.; Van Der Kraak, G.; Sorensen, P.W. Ten-week exposure to treated sewage discharge has relatively minor, variable effects on reproductive behavior and sperm production in goldfish. Environ. Toxicol. Chem. 2002, 21, 2185–2190. [Google Scholar] [CrossRef] [PubMed]
- McAllister, B.G.; Kime, D.E. Early life exposure to environmental levels of the aromatase inhibitor tributyltin causes masculinisation and irreversible sperm damage in zebrafish (Danio rerio). Aquat. Toxicol. 2003, 65, 309–316. [Google Scholar] [CrossRef]
- Haubruge, E.; Petit, F.; Gage, M.J.G. Reduced sperm counts in guppies (Poecilia reticulata) following exposure to low levels of tributyltin and bisphenol A. Proc. Biol. Sci. 2000, 267, 2333–2337. [Google Scholar] [CrossRef] [Green Version]
- Hatef, A.; Alavi, S.M.H.; Abdulfatah, A.; Fontaine, P.; Rodina, M.; Linhart, O. Adverse effects of bisphenol A on reproductive physiology in male goldfish at environmentally relevant concentrations. Ecotoxicol. Environ. Saf. 2012, 76, 56–62. [Google Scholar] [CrossRef]
- Hatef, A.; Zare, A.; Alavi, S.M.H.; Habibi, H.R.; Linhart, O. Modulations of androgen and estrogen mediating genes and testicular response in male goldfish exposed to bisphenol A. Environ. Toxicol. Chem. 2012, 31, 2069–2077. [Google Scholar] [CrossRef]
- Golshan, M.; Hatef, A.; Socha, M.; Milla, S.; Butts, I.A.E.; Carnevali, O.; Rodina, M.; Sokołowska-Mikołajczyk, M.; Fontaine, P.; Linhart, O.; et al. Di-(2-ethylhexyl)-phthalate disrupts pituitary and testicular hormonal functions to reduce sperm quality in mature goldfish. Aquat. Toxicol. 2015, 163, 16–26. [Google Scholar] [CrossRef]
- Baatrup, E.; Junge, M. Antiandrogenic pesticides disrupt sexual characteristics in the adult male guppy (Poecilia reticulata). Environ. Health Perspect. 2001, 109, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Bayley, M.; Foged Larsen, P.; Bækgaard, H.; Baatrup, E. The effects of vinclozolin, an anti-androgenic fungicide, on male guppy secondary sex characters and reproductive success. Biol. Reprod. 2003, 69, 1951–1956. [Google Scholar] [CrossRef] [Green Version]
- Hatef, A.; Alavi, S.M.H.; Milla, S.; Křišťan, J.; Golshan, M.; Fontaine, P.; Linhart, O. Anti-androgen vinclozolin impairs sperm quality and steroidogenesis in goldfish. Aquat. Toxicol. 2012, 122–123, 181–187. [Google Scholar] [CrossRef]
- Yin, P.; Li, Y.W.; Chen, Q.L.; Liu, Z.H. Diethylstilbestrol, flutamide and their combination impaired the spermatogenesis of male adult zebrafish through disrupting HPG axis, meiosis and apoptosis. Aquat. Toxicol. 2017, 185, 129–137. [Google Scholar] [CrossRef]
- Runnalls, T.; Hala, D.N.; Sumpter, J.P. Preliminary studies into the effects of the human pharamecutical Clofibtric acid on sperm parameters in adult fathead minnows. Aquat. Toxicol. 2007, 84, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Mennigen, J.A.; Lado, W.E.; Zamora, J.M.; Duarte-Guterman, P.; Langlois, V.S.; Metcalfe, C.D.; Chang, J.P.; Moon, T.W.; Trudeau, V.L. Waterborne fluoxetine disrupts the reproductive axis in sexually mature male goldfish, Carassius auratus. Aquat. Toxicol. 2010, 100, 354–364. [Google Scholar] [CrossRef]
- Roggio, M.A.; Guyon, N.F.; Hued, A.C.; Ame, M.V.; Valdes, M.E.; Giojalas, L.C.; Wunderlin, D.A.; Bistoni, M.A. Effects of the synthetic estrogen 17α-ethinylestradiol on aromatase expression, reproductive behavior and sperm quality in the fish Jenynsia multidentata. Bull. Environ. Contam. Toxicol. 2014, 92, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Bertram, M.G.; Ecker, T.E.; Wong, B.B.M.; O’Bryan, M.K.; Baumgartner, J.B.; Martin, J.M.; Saaristo, M. The antidepressant fluoxetine alters mechanisms of pre- and post-copulatory sexual selection in the eastern mosquitofish (Gambusia holbrooki). Environ. Pollut. 2018, 238, 238–247. [Google Scholar] [CrossRef]
- Sayed, A.E.D.H.; Mahmoud, U.M.; Mekkawy, I.A. Reproductive biomarkers to identify endocrine disruption in Clarias gariepinus exposed to 4-nonylphenol. Ecotoxicol. Environ. Saf. 2012, 78, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Franco, J.G., Jr.; Baruffi, R.L.; Mauri, A.L.; Petersen, C.G.; Oliveira, J.B.; Vagnini, L. Significance of large nuclear vacuoles in human spermatozoa: Implications for ICSI. Reprod. Biomed. Online 2008, 17, 42–45. [Google Scholar] [CrossRef]
- Yoshikawa, H.; Morishima, K.; Fujimoto, T.; Arias-Rodriguez, L.; Yamaha, E.; Arai, K. Ploidy manipulation using diploid sperm in the loach, Misgurnus anguillicaudatus: A review. J. Appl. Ichthyol. 2008, 24, 410–414. [Google Scholar] [CrossRef] [Green Version]
- Na-Nakorn, U.; Rangsin, W.; Boonngam, J. Allotriploidy increases sterility in the hybrid between Clarias macrocephalus and Clarias gariepinus. Aquaculture 2004, 237, 73–88. [Google Scholar] [CrossRef]
- Brown, K.H.; Schultz, I.R.; Cloud, J.G.; Nagler, J.J. Aneuploid sperm formation in rainbow trout exposed to the environmental estrogen 17α-ethynylestradiol. Proc. Natl. Acad. Sci. USA 2008, 105, 19786–19791. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yuan, C.; Chen, S.; Zheng, Y.; Zhang, Y.; Gao, J.; Wang, Z. Global and cyp19a1a gene specific DNA methylation in gonads of adult rare minnow Gobiocypris rarus under Bisphenol A exposure. Aquat. Toxicol. 2014, 156, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.; Zhang, Y.; Liu, Y.; Zhang, T.; Wang, Z. Enhanced GSH synthesis by Bisphenol A exposure promoted DNA methylation process in the testes of adult rare minnow Gobiocypris rarus. Aquat. Toxicol. 2016, 178, 99–105. [Google Scholar] [CrossRef]
- Horii, T.; Hatada, I. Regulation of CpG methylation by Dnmt and Tet in pluripotent stem cells. J. Reprod. Dev. 2016, 62, 331–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frankel, T.; Yonkos, L.; Ampy, F.; Frankel, J. Exposure to levonorgestrel increases nest acquisition success and decreases sperm motility in the male fathead minnow (Pimephales promelas). Environ. Toxicol. Chem. 2018, 37, 1131–1137. [Google Scholar] [CrossRef]
- Zebral, Y.D.; Anni, I.S.A.; Junior, A.S.V.; Corcini, C.D.; da Silva, J.C.; Caldas, J.S.; Acosta, I.B.; Afonso, S.B.; Bianchini, A. Life-time exposure to waterborne copper IV: Sperm quality parameters are negatively affected in the killifish Poecilia vivipara. Chemosphere 2019, 236, 124332. [Google Scholar] [CrossRef]
- Murack, P.J.; Parrish, J.; Barry, T.P. Effects of progesterone on sperm motility in fathead minnow (Pimephales promelas). Aquat. Toxicol. 2011, 104, 121–125. [Google Scholar] [CrossRef]
- Das Neves, J.; Barnhoorn, I.E.J.; Wagenaar, G.M. The effects of environmentally relevant concentrations of aldrin and methoxychlor on the testes and sperm of male Clarias gariepinus (Burchell, 1822) after short-term exposure. Fish Physiol. Biochem. 2018, 44, 1421–1434. [Google Scholar] [CrossRef]
- Martin, J.M.; Bertram, M.G.; Saaristo, M.; Ecker, T.E.; Hannington, S.L.; Tanner, J.L.; Michelangeli, M.; O’Bryan, M.K.; Wong, B.B.M. Impact of the widespread pharmaceutical pollutant fluoxetine on behaviour and sperm traits in a freshwater fish. Sci. Total Environ. 2019, 650, 1771–1778. [Google Scholar] [CrossRef]
- Guyón, N.F.; Roggio, M.A.; Amé, M.V.; Hued, A.C.; Valdés, M.E.; Giojalas, L.C.; Wunderlin, D.A.; Bistoni, M.A. Impairments in aromatase expression, reproductive behavior, and sperm quality of male fish exposed to 17β-estradiol. Environ. Toxicol. Chem. 2012, 31, 935–940. [Google Scholar] [CrossRef]
- Soares, J.; Coimbra, A.M.; Reis-Henriques, M.A.; Monteiro, N.M.; Vieira, M.N.; Oliveira, J.M.A.; Guedes-Dias, P.; Fontaínhas-Fernandes, A.; Silva Parra, S.; Carvalho, A.P.; et al. Disruption of zebrafish (Danio rerio) embryonic development after full life-cycle parental exposure to low levels of ethinylestradiol. Aquat. Toxicol. 2009, 95, 330–338. [Google Scholar] [CrossRef]
- Velicu, M.; Suri, R. Presence of steroid hormones and antibiotics in surface water of agricultural, suburban and mixed-use areas. Environ. Monit. Assess. 2009, 154, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Kuch, H.M.; Ballschmitter, K. Determination of endocrine-disrupting phenolic compounds and estrogens in surface and drinking water by HRGC-(NCI)-MS in the picogram per liter range. Environ. Sci. Technol. 2001, 35, 3201–3206. [Google Scholar] [CrossRef]
- Baronti, C.; Curini, R.; D’Ascenzo, G.; Di Corcia, A.; Gentili, A.; Samperi, R. Monitoring natural and synthetic estrogens at activated treatment plants and in receiving river water. Environ. Sci. Technol. 2000, 34, 5059–5066. [Google Scholar] [CrossRef]
- Nasu, M.; Goto, M.; Kato, H.; Oshima, Y.; Tanaka, H. Study on endocrine disrupting chemicals in wastewater treatment plants. Water Sci. Technol. 2000, 43, 101–108. [Google Scholar] [CrossRef]
- Desbrow, C.; Routledge, E.J.; Brighty, G.C.; Sumpter, J.P.; Waldock, M. Identification of estrogenic chemicals in STW effluent: 1. Chemical fractionation and in vitro biological screening. Environ. Sci. Technol. 1998, 32, 1549–1558. [Google Scholar] [CrossRef]
- Liu, S.; Ying, G.G.; Zhao, J.L.; Chen, F.; Yang, B.; Zhou, L.J.; Lai, H.J. Trace analysis of 28 steroids in surface water, wastewater and sludge samples by rapid resolution liquid chromatography-electrospray ionization tandem mass spectrometry. J. Chromatogr. A 2011, 1218, 1367–1378. [Google Scholar] [CrossRef] [PubMed]
- Belfroid, A.C.; Van der Horst, A.; Vethaak, A.D.; Schafer, A.J.; Ris, G.B.J.; Wegener, J.; Cofino, W.P. Analysis and occurrence of estrogenic hormones and their glucuronides in surface water and waste water in the Netherlands. Sci. Total Environ. 1999, 225, 101–108. [Google Scholar] [CrossRef]
- Ternes, T.A.; Stumpf, M.; Mueller, J.; Haberer, K.; Wilken, R.D.; Servos, M. Behaviour and occurrence of estrogens in municipal sewage treatment plants—I. Investigations in Germany, Canada and Brazil. Sci. Total Environ. 1999, 225, 81–90. [Google Scholar] [CrossRef]
- Snyder, S.; Giesy, J.; Kannan, K.; Gross, T.; Snyder, E.; Verbrugge, D.; Keith, T. Analytical methods for detection of selected estrogenic compounds in aqueous mixtures. Environ. Sci. Technol. 1999, 33, 2814–2820. [Google Scholar] [CrossRef]
- Yu, Q.; Geng, J.; Ren, H. Occurrence and fate of androgens in municipal wastewater treatment plants in China. Chemosphere 2019, 237, 124371. [Google Scholar] [CrossRef] [PubMed]
- Blankvoort, B.M.G.; Rodenburg, R.J.T.; Murk, A.J.; Koeman, J.H.; Schilt, R.; Aarts, J.M.M.J.G. Androgenic activity in surface water samples detected using the AR-LUX assay: Indication for mixture effects. Environ. Toxicol. Pharmacol. 2005, 19, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Höhne, C.; Püttmann, W. Occurrence and temporal variations of the xenoestrogens bisphenol A, 4-tert-octylphenol, and tech. 4-nonylphenol in two German wastewater treatment plants. Environ. Sci. Pollut. Res. Int. 2008, 15, 405–416. [Google Scholar] [CrossRef]
- Cailleaud, K.; Forget-Leray, J.; Souissi, S.; Lardy, S.; Augagneur, S.; Budzinski, H. Seasonal variation of hydrophobic organic contaminant concentrations in the water-column of the Seine Estuary and their transfer to a planktonic species Eurytemora affinis (Calanoid, copepod). Part 2: Alkylphenol-polyethoxylates. Chemosphere 2007, 70, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, S.; Huang, C.H.; Mukherji, S.; Padhye, L.P. Occurrence and fate of pharmaceuticals in WWTPs in India and comparison with a similar study in the United States. Chemosphere 2016, 159, 526–535. [Google Scholar] [CrossRef]
- Kang, J.H.; Aasi, D.; Katayama, Y. Bisphenol A in the aquatic environment and its endocrine-disruptive effects on aquatic organisms. Crit. Rev. Toxicol. 2007, 37, 607–625. [Google Scholar] [CrossRef]
- Fromme, H.; Küchler, T.; Otto, T.; Pilz, K.; Müller, J.; Wenzel, A. Occurrence of phthalates and bisphenol A and F in the environment. Water Res. 2002, 36, 1429–1438. [Google Scholar] [CrossRef]
- Yang, Y.; Lu, L.; Zhang, J.; Yang, Y.; Wu, Y.; Shao, B. Simultaneous determination of seven bisphenols in environmental water and solid samples by liquid chromatography-electrospray tandem mass spectrometry. J. Chromatogr. A 2014, 1328, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Bókony, V.; Üveges, B.; Ujhegyi, N.; Verebélyi, V.; Nemesházi, E.; Csíkvári, O.; Hettyey, A. Endocrine disruptors in breeding ponds and reproductive health of toads in agricultural, urban and natural landscapes. Sci. Total Environ. 2018, 634, 1335–1345. [Google Scholar] [CrossRef]
- Wen, Z.; Huang, X.; Gao, D.; Liu, G.; Fang, C.; Shang, Y.; Du, J.; Zhao, Y.; Lv, L.; Song, K. Phthalate esters in surface water of Songhua River watershed associated with land use types, Northeast China. Environ. Sci. Pollut. Res. Int. 2018, 25, 7688–7698. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.M.; Zhang, H.H.; Zou, Y.W.; Yang, G.P. Distribution and ecotoxicological state of phthalate esters in the sea-surface microlayer, seawater and sediment of the Bohai Sea and the Yellow Sea. Environ. Pollut. 2018, 240, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Readman, J.W.; Albanis, T.A.; Barcelo, D.; Galassi, S.; Tronczynski, J.; Gabrielides, G.P. Fungicide contamination of Mediterranean estuarine waters: Results from a MED POL pilot survey. Mar. Poll. Bull. 1997, 34, 259–263. [Google Scholar] [CrossRef]
- Kreuger, J.; Graaf, S.; Patring, J.; Adielsson, S. Pesticides in Surface Water in Areas with Open Ground and Greenhouse Horticultural Crops in Sweden 2008; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2010. [Google Scholar]
- Zheng, S.; Chen, B.; Qiu, X.; Chen, M.; Ma, Z.; Yu, X. Distribution and risk assessment of 82 pesticides in Jiulong River and estuary in South China. Chemosphere 2016, 144, 1177–1192. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ying, G.G.; Zhao, J.L.; Liu, S.; Yang, B.; Zhou, L.J.; Tao, R.; Su, H.C. Assessing estrogenic activity in surface water and sediment of the Liao River system in northeast China using combined chemical and biological tools. Environ. Pollut. 2011, 159, 148–156. [Google Scholar] [CrossRef]
- Chen, T.S.; Chen, T.C.; Yeh, K.J.C.; Chao, H.R.; Liaw, E.T.; Hsieh, C.Y.; Chen, K.C.; Hsieh, L.T.; Yeh, Y.L. High estrogen concentrations in receiving river discharge from a concentrated livestock feedlot. Sci. Total Environ. 2010, 408, 3223–3230. [Google Scholar] [CrossRef] [PubMed]
- Lei, B.; Huang, S.; Zhou, Y.; Wang, D.; Wang, Z. Levels of six estrogens in water and sediment from three rivers in Tianjin area, China. Chemosphere 2009, 76, 36–42. [Google Scholar] [CrossRef]
- Pojana, G.; Gomiero, A.; Jonkers, N.; Marcomini, A. Natural and synthetic endocrine disrupting compounds (EDCs) in water, sediment and biota of a coastal lagoon. Environ. Int. 2007, 33, 929–936. [Google Scholar] [CrossRef]
- Wu, M.; Xiang, J.; Que, C.; Chen, F.; Xu, G. Occurrence and fate of psychiatric pharmaceuticals in the urban water system of Shanghai, China. Chemosphere 2015, 138, 486–493. [Google Scholar] [CrossRef]
- Paíga, P.; Santos, L.H.; Ramos, S.; Jorge, S.; Silva, J.G.; Delerue-Matos, C. Presence of pharmaceuticals in the Lis river (Portugal): Sources, fate and seasonal variation. Sci. Total Environ. 2016, 573, 164–177. [Google Scholar] [CrossRef]
- Lindim, C.; van Gils, J.; Georgieva, D.; Mekenyan, O.; Cousins, I.T. Evaluation of human pharmaceutical emissions and concentrations in Swedish river basins. Sci. Total Environ. 2016, 572, 508–519. [Google Scholar] [CrossRef]
- Santos, L.H.; Gros, M.; Rodriguez-Mozaz, S.; Delerue-Matos, C.; Pena, A.; Barceló, D.; Montenegro, M.C.B. Contribution of hospital effluents to the load of pharmaceuticals in urban wastewaters: Identification of ecologically relevant pharmaceuticals. Sci. Total Environ. 2013, 461, 302–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Q.; Zhang, Y.X.; Kang, J.; Gan, X.M.; Xu, Y.P.; Guo, J.S.; Gao, X. A preliminary study on the occurrence of pharmaceutically active compounds in the river basins and their removal in two conventional drinking water treatment plants in Chongqing, China. Clean Soil Air Water 2015, 43, 794–803. [Google Scholar] [CrossRef]
- Nentwig, G.; Oetken, M.; Oehlmann, J. Effects of Pharmaceuticals on Aquatic Invertebrates—The Example of Carbamazepine and Clofibric Acid. In Pharmaceuticals in the Environment. Sources, Fate, Effects and Risks; Kümmerer, K., Ed.; Springer: Heidelberg, Germany, 2004; pp. 195–208. [Google Scholar] [CrossRef] [Green Version]
- Anjum, R.; Malik, A. Evaluation of mutagenicity of wastewater in the vicinity of pesticide industry. Environ. Toxicol. Pharmacol. 2013, 35, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Zhang, G. Analysis of organophosphorous pesticides from source water using solid-phase extraction technique. China Environ. Sci. 2000, 20, 1–4. [Google Scholar]
- Kumari, B.; Madan, V.K.; Kathpal, T.S. Pesticide residues in rain water from Hisar, India. Environ. Monit. Assess. 2007, 133, 467–471. [Google Scholar] [CrossRef]
- Canty, M.N.; Hutchinson, T.H.; Brown, R.J.; Jones, M.B.; Jha, A.N. Linking genotoxic responses with cytotoxic and behavioural or physiological consequences: Differential sensitivity of echinoderms (Asterias rubens) and marine molluscs (Mytilus edulis). Aquat. Toxicol. 2009, 94, 68–76. [Google Scholar] [CrossRef]
- Guo, S.; Qian, L.; Shi, H.; Barry, T.; Cao, Q.; Liu, J. Effects of tributyltin (TBT) on Xenopus tropicalis embryos at environmentally relevant concentrations. Chemosphere 2010, 79, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yin, P.; Zhao, L. Phthalate esters in water and surface sediments of the Pearl River Estuary: Distribution, ecological, and human health risks. Environ. Sci. Pollut. Res. 2016, 23, 19341–19349. [Google Scholar] [CrossRef]
- Nibamureke, U.M.C.; Barnhoorn, I.E.J.; Wagenaar, G.M. Health assessment of freshwater fish species from Albasini Dam, outside a DDT-sprayed area in Limpopo province, South Africa: A preliminary study. Afr. J. Aquat. Sci. 2016, 41, 1–12. [Google Scholar] [CrossRef]
- Song, S.; Ruan, T.; Wang, T.; Liu, R.; Jiang, G. Distribution and preliminary exposure assessment of bisphenol AF (BPAF) in various environmental matrices around a manufacturing plant in China. Environ. Sci. Technol. 2012, 46, 13136–13143. [Google Scholar] [CrossRef]
- Corcoran, J.; Winter, M.J.; Tyler, C.R. Pharmaceuticals in the aquatic environment: A critical review of the evidence for health effects in fish. Crit. Rev. Toxicol. 2010, 40, 287–304. [Google Scholar] [CrossRef]
- Patel, M.; Kumar, R.; Kishor, K.; Mlsna, T.; Pittman, C.U., Jr.; Mohan, D. Pharmaceuticals of emerging concern in aquatic systems: Chemistry, occurrence, effects, and removal methods. Chem. Rev. 2019, 119, 3510–3673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dsikowitzky, L.; Schwarzbauer, J. Industrial organic contaminants: Identification, toxicity and fate in the environment. Environ. Chem. Lett. 2014, 12, 371–386. [Google Scholar] [CrossRef]
- Petrie, B.; Barden, R.; Kasprzyk-Hordern, B. A review on emerging contaminants in wastewaters and the environment: Current knowledge, understudied areas and recommendations for future monitoring. Water Res. 2015, 72, 3–27. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, R.P.; Egli, T.; Hofstetter, T.B.; von Gunten, U.; Wehrli, B. Global water pollution and human health. Ann. Rev. Environ. Resour. 2010, 35, 109–136. [Google Scholar] [CrossRef]
- Denslow, N.; Sepulveda, M. Ecotoxicological effects of endocrine disrupting compounds on fish reproduction. In The Fish Oocyte: From Basic Studies to Biotechnological Applications; Babin, P.J., Cerdà, J., Lubzens, E., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 255–322. [Google Scholar] [CrossRef]
- Dietrich, M.A.; Nynca, J.; Ciereszko, A. Proteomic and metabolomic insights into the functions of the male reproductive system in fishes. Theriogenology 2019, 132, 182–200. [Google Scholar] [CrossRef]
- Hatef, A.; Alavi, S.M.H.; Butts, I.A.E.; Policar, T.; Linhart, O. The mechanisms of action of mercury on sperm morphology, adenosine-5′-triphosphate content and motility in Perca fluviatilis (Percidae; Teleostei). Environ. Toxicol. Chem. 2011, 30, 905–914. [Google Scholar] [CrossRef]
- Schiffer, C.; Müller, A.; Egeberg, D.L.; Alvarez, L.; Brenker, C.; Rehfeld, A.; Frederiksen, H.; Wäschle, B.; Kaupp, U.B.; Balbach, M.; et al. Direct action of endocrine disrupting chemicals on human sperm. EMBO Rep. 2014, 15, 758–765. [Google Scholar] [CrossRef] [Green Version]
- De la Fuente, I.M.; Cortés, J.M.; Valero, E.; Desroches, M.; Rodrigues, S.; Malaina, I.; Martínez, L. On the dynamics of the adenylate energy system: Homeorhesis vs. homeostasis. PLoS ONE 2014, 9, e108676. [Google Scholar] [CrossRef] [Green Version]
- Mills, L.J.; Chichester, C. Review of evidence: Are endocrine-disrupting chemicals in the aquatic environment impacting fish populations? Sci. Total Environ. 2005, 343, 1–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Zhu, X.S.; Cai, Z.H. Endocrine disruptors: An overview and discussion on issues surrounding their impact on marine animals. J. Mar. Anim. Ecol. 2009, 2, 7–17. [Google Scholar]
- Hamilton, P.B.; Cowx, I.G.; Oleksiak, M.F.; Griffiths, A.M.; Grahn, M.; Stevens, J.R.; Carvalho, G.R.; Nicol, E.; Tyler, C.R. Population-level consequences for wild fish exposed to sublethal concentrations of chemicals—A critical review. Fish Fish. 2016, 17, 545–566. [Google Scholar] [CrossRef]
- Boryshpolets, S.; Kowalski, R.K.; Dietrich, G.J.; Dzyuba, B.; Ciereszko, A. Different computer-assisted sperm analysis (CASA) systems highly influence sperm motility parameters. Theriogenology 2013, 80, 758–765. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alavi, S.M.H.; Barzegar-Fallah, S.; Rahdar, P.; Ahmadi, M.M.; Yavari, M.; Hatef, A.; Golshan, M.; Linhart, O. A Review on Environmental Contaminants-Related Fertility Threat in Male Fishes: Effects and Possible Mechanisms of Action Learned from Wildlife and Laboratory Studies. Animals 2021, 11, 2817. https://doi.org/10.3390/ani11102817
Alavi SMH, Barzegar-Fallah S, Rahdar P, Ahmadi MM, Yavari M, Hatef A, Golshan M, Linhart O. A Review on Environmental Contaminants-Related Fertility Threat in Male Fishes: Effects and Possible Mechanisms of Action Learned from Wildlife and Laboratory Studies. Animals. 2021; 11(10):2817. https://doi.org/10.3390/ani11102817
Chicago/Turabian StyleAlavi, Sayyed Mohammad Hadi, Sepideh Barzegar-Fallah, Parastoo Rahdar, Mohammad Mahdi Ahmadi, Mina Yavari, Azadeh Hatef, Mahdi Golshan, and Otomar Linhart. 2021. "A Review on Environmental Contaminants-Related Fertility Threat in Male Fishes: Effects and Possible Mechanisms of Action Learned from Wildlife and Laboratory Studies" Animals 11, no. 10: 2817. https://doi.org/10.3390/ani11102817