

Mosquito Salivary Proteins and Arbovirus Infection: From Viral Enhancers to Potential Targets for Vaccines
 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Emerging Arboviruses, an Increased Concern for Public Health
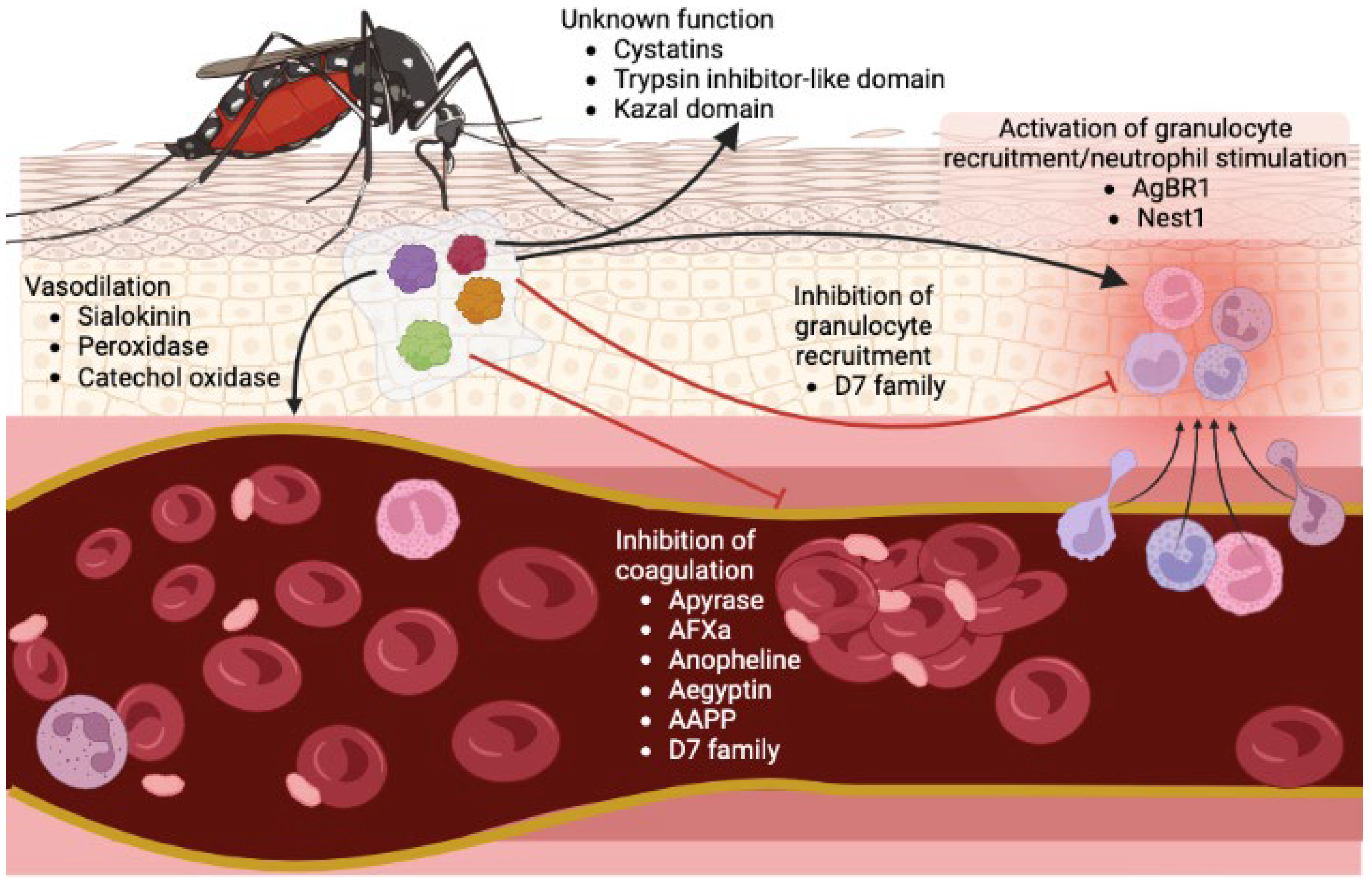
2. Salivary Proteins Facilitate Mosquito Blood Feeding
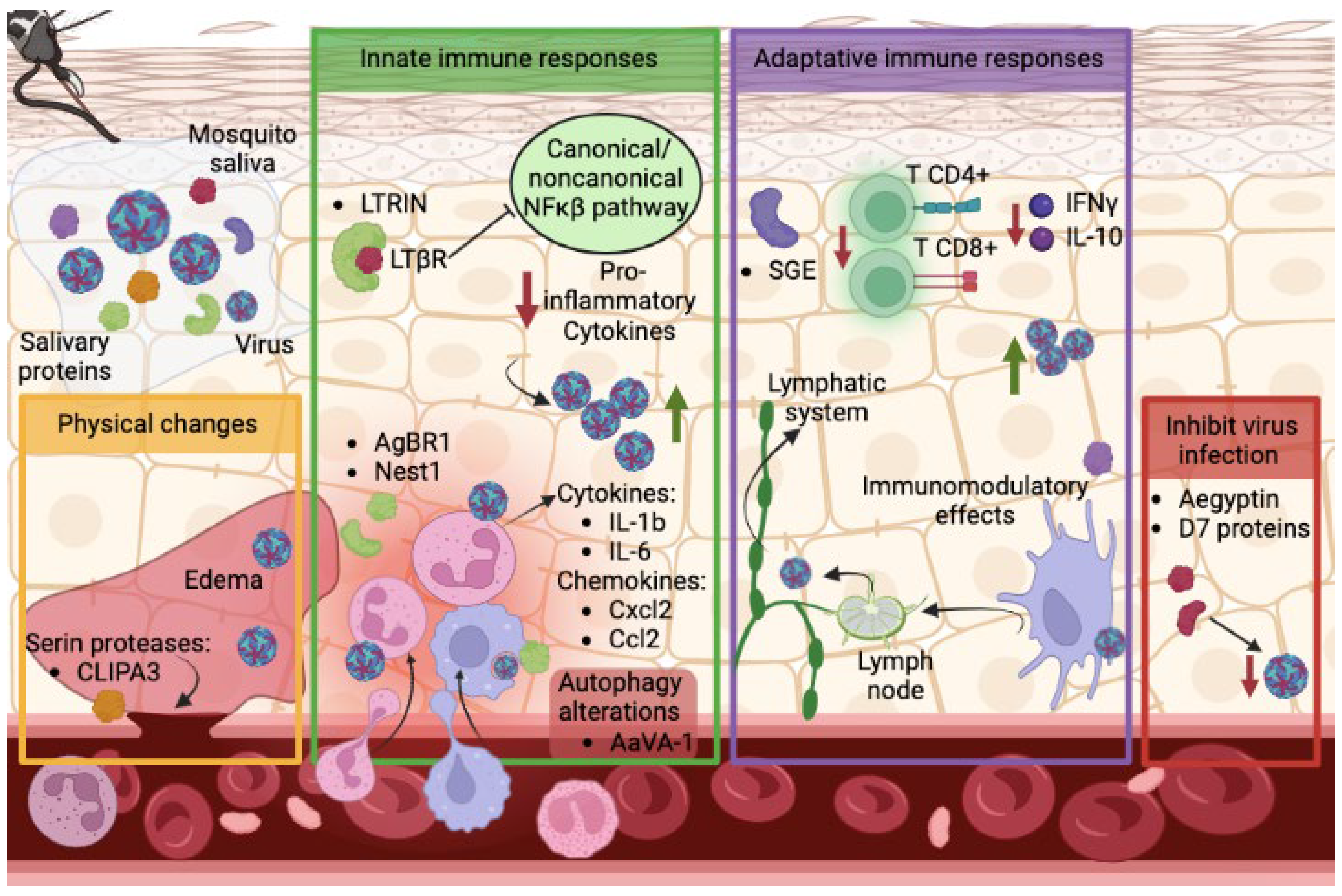
3. Mosquito Salivary Proteins Can Alter the Course of Arbovirus Dissemination and Transmission
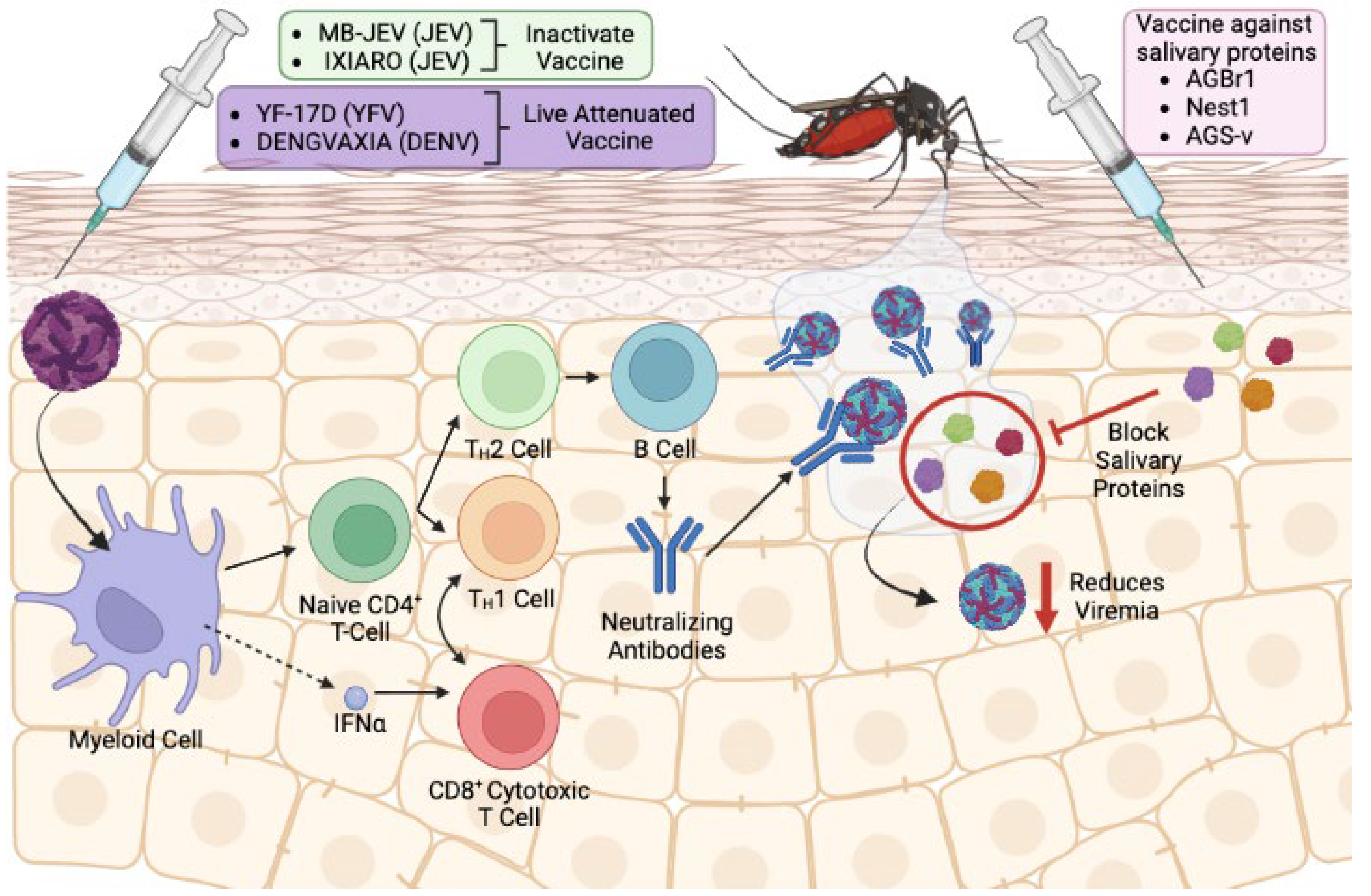
4. The Use of Salivary Proteins as Vector-Based Vaccines: Benefits and Potential Pitfalls
5. Immune Responses to Saliva Proteins as Biomarkers of Exposure
6. Conclusions and Future Challenges
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Campbell, L.P.; Luther, C.; Moo-Llanes, D.; Ramsey, J.M.; Danis-Lozano, R.; Peterson, A.T. Climate change influences on global distributions of dengue and chikungunya virus vectors. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140135. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.G.; Harris, E. Dengue. Lancet 2015, 385, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J. Dengue, Urbanization and Globalization: The Unholy Trinity of the 21(st) Century. Trop. Med. Health 2011, 39 (Suppl. S4), 3–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Alwis, R.; Beltramello, M.; Messer, W.B.; Sukupolvi-Petty, S.; Wahala, W.M.; Kraus, A.; Olivarez, N.P.; Pham, Q.; Brien, J.D.; Tsai, W.Y.; et al. In-depth analysis of the antibody response of individuals exposed to primary dengue virus infection. PLoS. Negl. Trop. Dis. 2011, 5, e1188. [Google Scholar] [CrossRef]
- Macnamara, F.N. Zika virus: A report on three cases of human infection during an epidemic of jaundice in Nigeria. Trans. R. Soc. Trop. Med. Hyg. 1954, 48, 139–145. [Google Scholar] [CrossRef]
- Duffy, M.R.; Chen, T.H.; Hancock, W.T.; Powers, A.M.; Kool, J.L.; Lanciotti, R.S.; Pretrick, M.; Marfel, M.; Holzbauer, S.; Dubray, C.; et al. Zika virus outbreak on Yap Island, Federated States of Micronesia. N. Engl. J. Med. 2009, 360, 2536–2543. [Google Scholar] [CrossRef]
- Campos, G.S.; Bandeira, A.C.; Sardi, S.I. Zika Virus Outbreak, Bahia, Brazil. Emerg. Infect. Dis. 2015, 21, 1885–1886. [Google Scholar] [CrossRef]
- Enfissi, A.; Codrington, J.; Roosblad, J.; Kazanji, M.; Rousset, D. Zika virus genome from the Americas. Lancet 2016, 387, 227–228. [Google Scholar] [CrossRef] [Green Version]
- Hennessey, M.; Fischer, M.; Staples, J.E. Zika Virus Spreads to New Areas-Region of the Americas, May 2015-January 2016. Morb. Mortal. Wkly. Rep. 2016, 65, 55–58. [Google Scholar] [CrossRef]
- May, F.J.; Davis, C.T.; Tesh, R.B.; Barrett, A.D. Phylogeography of West Nile virus: From the cradle of evolution in Africa to Eurasia, Australia, and the Americas. J. Virol. 2011, 85, 2964–2974. [Google Scholar] [CrossRef] [Green Version]
- Debiasi, R.L.; Tyler, K.L. West Nile virus meningoencephalitis. Nat. Clin. Pract. Neurol. 2006, 2, 264–275. [Google Scholar] [CrossRef]
- Granwehr, B.P.; Lillibridge, K.M.; Higgs, S.; Mason, P.W.; Aronson, J.F.; Campbell, G.A.; Barrett, A.D. West Nile virus: Where are we now? Lancet Infect. Dis. 2004, 4, 547–556. [Google Scholar] [CrossRef]
- Ronca, S.E.; Ruff, J.C.; Murray, K.O. A 20-year historical review of West Nile virus since its initial emergence in North America: Has West Nile virus become a neglected tropical disease? PLoS Negl. Trop. Dis. 2021, 15, e0009190. [Google Scholar] [CrossRef]
- Ribeiro, J.M. Characterization of a vasodilator from the salivary glands of the yellow fever mosquito Aedes aegypti. J. Exp. Biol. 1992, 165, 61–71. [Google Scholar] [CrossRef]
- Ribeiro, J.M.C. Insect Saliva: Function, Biochemistry, and Physiology. In Regulatory Mechanisms in Insect Feeding; Chapman, R.F., de Boer, G., Eds.; Springer: Boston, MA, USA, 1995; pp. 74–97. [Google Scholar]
- Ribeiro, J.M.C.; Mans, B.J.; Arcà, B. An insight into the sialome of blood-feeding Nematocera. Insect Biochem. Mol. Biol. 2010, 40, 767–784. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, J.M.; Arca, B.; Lombardo, F.; Calvo, E.; Phan, V.M.; Chandra, P.K.; Wikel, S.K. An annotated catalogue of salivary gland transcripts in the adult female mosquito, Aedes aegypti. BMC Genom. 2007, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela, J.G.; Pham, V.M.; Garfield, M.K.; Francischetti, I.M.B.; Ribeiro, J.M.C. Toward a description of the sialome of the adult female mosquito Aedes aegypti. Insect Biochem. Mol. Biol. 2002, 32, 1101–1122. [Google Scholar] [CrossRef]
- Arcà, B.; Lombardo, F.; Francischetti, I.M.B.; Pham, V.M.; Mestres-Simon, M.; Andersen, J.F.; Ribeiro, J.M.C. An insight into the sialome of the adult female mosquito Aedes albopictus. Insect Biochem. Mol. Biol. 2007, 37, 107–127. [Google Scholar] [CrossRef]
- Ribeiro, J.M.C.; Charlab, R.; Pham, V.M.; Garfield, M.; Valenzuela, J.G. An insight into the salivary transcriptome and proteome of the adult female mosquito Culex pipiens quinquefasciatus. Insect Biochem. Mol. Biol. 2004, 34, 543–563. [Google Scholar] [CrossRef]
- Arcà, B.; Lombardo, F.; de Lara Capurro, M.; della Torre, A.; Dimopoulos, G.; James, A.A.; Coluzzi, M. Trapping cDNAs encoding secreted proteins from the salivary glands of the malaria vector Anopheles gambiae. Proc. Natl. Acad. Sci. USA 1999, 96, 1516–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.; McNicol, A.; James, A.A.; Peng, Z. Expression of functional recombinant mosquito salivary apyrase: A potential therapeutic platelet aggregation inhibitor. Platelets 2006, 17, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.M. Role of saliva in blood-feeding by arthropods. Annu. Rev. Entomol. 1987, 32, 463–478. [Google Scholar] [CrossRef] [PubMed]
- Smartt, C.T.; Kim, A.P.; Grossman, G.L.; James, A.A. The Apyrase Gene of the Vector Mosquito, Aedes eegypti, Is Expressed Specifically in the Adult Female Salivary Glands. Exp. Parasitol. 1995, 81, 239–248. [Google Scholar] [CrossRef]
- Ribeiro, J.M.C. Blood-feeding in mosquitoes: Probing time and salivary gland anti-haemostatic activities in representatives of three genera (Aedes, Anopheles, Culex). Med. Vet. Entomol. 2000, 14, 142–148. [Google Scholar] [CrossRef]
- Ribeiro, J.M.C.; Valenzuela, J.G. The salivary purine nucleosidase of the mosquito, Aedes aegypti. Insect Biochem. Mol. Biol. 2003, 33, 13–22. [Google Scholar] [CrossRef]
- Tilley, S.L.; Wagoner, V.A.; Salvatore, C.A.; Jacobson, M.A.; Koller, B.H. Adenosine and inosine increase cutaneous vasopermeability by activating A3 receptors on mast cells. J. Clin. Investig. 2000, 105, 361–367. [Google Scholar] [CrossRef] [Green Version]
- Calvo, E.; Ribeiro, J.M.C. A novel secreted endonuclease from Culex quinquefasciatussalivary glands. J. Exp. Biol. 2006, 209, 2651–2659. [Google Scholar] [CrossRef] [Green Version]
- Argentine, J.A.; James, A.A. Characterization of a salivary gland-specific esterase in the vector mosquito, Aedes aegypti. Insect Biochem. Mol. Biol. 1995, 25, 621–630. [Google Scholar] [CrossRef]
- Champagne, D.E.; Ribeiro, J.M. Sialokinin I and II: Vasodilatory tachykinins from the yellow fever mosquito Aedes aegypti. Proc. Natl. Acad. Sci USA 1994, 91, 138–142. [Google Scholar] [CrossRef] [Green Version]
- Beerntsen, B.T.; Champagne, D.E.; Coleman, J.L.; Campos, Y.A.; James, A.A. Characterization of the Sialokinin I gene encoding the salivary vasodilator of the yellow fever mosquito, Aedes aegypti. Insect Mol. Biol. 1999, 8, 459–467. [Google Scholar] [CrossRef]
- Martin-Martin, I.; Valenzuela Leon, P.C.; Amo, L.; Shrivastava, G.; Iniguez, E.; Aryan, A.; Brooks, S.; Kojin, B.B.; Williams, A.E.; Bolland, S.; et al. Aedes aegypti sialokinin facilitates mosquito blood feeding and modulates host immunity and vascular biology. Cell Rep. 2022, 39, 110648. [Google Scholar] [CrossRef]
- Ribeiro, J.M.; Nussenzveig, R.H. The salivary catechol oxidase/peroxidase activities of the mosquito Anopheles albimanus. J. Exp. Biol. 1993, 179, 273–287. [Google Scholar] [CrossRef]
- Ribeiro, J.M.; Valenzuela, J.G. Purification and cloning of the salivary peroxidase/catechol oxidase of the mosquito Anopheles albimanus. J. Exp. Biol. 1999, 202, 809–816. [Google Scholar] [CrossRef]
- Stark, K.R.; James, A.A. A Factor Xa-Directed Anticoagulant from the Salivary Glands of the Yellow Fever Mosquito Aedes aegypti. Exp. Parasitol. 1995, 81, 321–331. [Google Scholar] [CrossRef]
- Stark, K.R.; James, A.A. Isolation and Characterization of the Gene Encoding a Novel Factor Xa-directed Anticoagulant from the Yellow Fever Mosquito, Aedes aegypti *. J. Biol. Chem. 1998, 273, 20802–20809. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, A.C.; de Sanctis, D.; Gutiérrez-Gallego, R.; Cereija, T.B.; Macedo-Ribeiro, S.; Fuentes-Prior, P.; Pereira, P.J.B. Unique thrombin inhibition mechanism by anophelin, an anticoagulant from the malaria vector. Proc. Natl. Acad. Sci. USA 2012, 109, E3649–E3658. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, K.; Walvekar, V.A.; Wong, B.; Sayed, A.M.M.; Missé, D.; Kini, R.M.; Mok, Y.K.; Pompon, J. Increased Mosquito Midgut Infection by Dengue Virus Recruitment of Plasmin Is Blocked by an Endogenous Kazal-type Inhibitor. iScience 2019, 21, 564–576. [Google Scholar] [CrossRef] [Green Version]
- James, A.A.; Blackmer, K.; Marinotti, O.; Ghosn, C.R.; Racioppi, J.V. Isolation and characterization of the gene expressing the major salivary gland protein of the female mosquito, Aedes aegypti. Mol. Biochem. Parasitol. 1991, 44, 245–253. [Google Scholar] [CrossRef]
- Valenzuela, J.G.; Charlab, R.; Gonzalez, E.C.; De Miranda-Santos, I.K.F.; Marinotti, O.; Francischetti, I.M.B.; Ribeiro, J.M.C. The D7 family of salivary proteins in blood sucking diptera. Insect Mol. Biol. 2002, 11, 149–155. [Google Scholar] [CrossRef]
- Calvo, E.; Mans, B.J.; Andersen, J.F.; Ribeiro, J.M.C. Function and Evolution of a Mosquito Salivary Protein Family*. J. Biol. Chem. 2006, 281, 1935–1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Martin, I.; Paige, A.; Valenzuela Leon, P.C.; Gittis, A.G.; Kern, O.; Bonilla, B.; Chagas, A.C.; Ganesan, S.; Smith, L.B.; Garboczi, D.N.; et al. ADP binding by the Culex quinquefasciatus mosquito D7 salivary protein enhances blood feeding on mammals. Nat. Commun. 2020, 11, 2911. [Google Scholar] [CrossRef] [PubMed]
- Martin-Martin, I.; Smith, L.B.; Chagas, A.C.; Sá-Nunes, A.; Shrivastava, G.; Valenzuela-Leon, P.C.; Calvo, E. Aedes albopictus D7 Salivary Protein Prevents Host Hemostasis and Inflammation. Biomolecules 2020, 10, 1372. [Google Scholar] [CrossRef] [PubMed]
- Isawa, H.; Yuda, M.; Orito, Y.; Chinzei, Y. A Mosquito Salivary Protein Inhibits Activation of the Plasma Contact System by Binding to Factor XII and High Molecular Weight Kininogen*. J. Biol. Chem. 2002, 277, 27651–27658. [Google Scholar] [CrossRef] [Green Version]
- Isawa, H.; Orito, Y.; Iwanaga, S.; Jingushi, N.; Morita, A.; Chinzei, Y.; Yuda, M. Identification and characterization of a new kallikrein-kinin system inhibitor from the salivary glands of the malaria vector mosquito Anopheles stephensi. Insect Biochem. Mol. Biol. 2007, 37, 466–477. [Google Scholar] [CrossRef]
- Calvo, E.; Tokumasu, F.; Marinotti, O.; Villeval, J.-L.; Ribeiro, J.M.C.; Francischetti, I.M.B. Aegyptin, a Novel Mosquito Salivary Gland Protein, Specifically Binds to Collagen and Prevents Its Interaction with Platelet Glycoprotein VI, Integrin α2β1, and von Willebrand Factor *. J. Biol. Chem. 2007, 282, 26928–26938. [Google Scholar] [CrossRef] [Green Version]
- Calvo, E.; Tokumasu, F.; Mizurini, D.M.; McPhie, P.; Narum, D.L.; Ribeiro, J.M.C.; Monteiro, R.Q.; Francischetti, I.M.B. Aegyptin displays high-affinity for the von Willebrand factor binding site (RGQOGVMGF) in collagen and inhibits carotid thrombus formation in vivo. FEBS J. 2010, 277, 413–427. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; Sudo, T.; Niimi, M.; Tao, L.; Sun, B.; Kambayashi, J.; Watanabe, H.; Luo, E.; Matsuoka, H. Inhibition of collagen-induced platelet aggregation by anopheline antiplatelet protein, a saliva protein from a malaria vector mosquito. Blood 2008, 111, 2007–2014. [Google Scholar] [CrossRef] [Green Version]
- Chagas, A.C.; Ramirez, J.L.; Jasinskiene, N.; James, A.A.; Ribeiro, J.M.; Marinotti, O.; Calvo, E. Collagen-binding protein, Aegyptin, regulates probing time and blood feeding success in the dengue vector mosquito, Aedes aegypti. Proc. Natl. Acad. Sci. USA 2014, 111, 6946–6951. [Google Scholar] [CrossRef] [Green Version]
- Islam, A.; Emran, T.B.; Yamamoto, D.S.; Iyori, M.; Amelia, F.; Yusuf, Y.; Yamaguchi, R.; Alam, M.S.; Silveira, H.; Yoshida, S. Anopheline antiplatelet protein from mosquito saliva regulates blood feeding behavior. Sci. Rep. 2019, 9, 3129. [Google Scholar] [CrossRef] [Green Version]
- Le Coupanec, A.; Babin, D.; Fiette, L.; Jouvion, G.; Ave, P.; Misse, D.; Bouloy, M.; Choumet, V. Aedes mosquito saliva modulates Rift Valley fever virus pathogenicity. PLoS Negl. Trop. Dis. 2013, 7, e2237. [Google Scholar] [CrossRef]
- Conway, M.J.; Watson, A.M.; Colpitts, T.M.; Dragovic, S.M.; Li, Z.; Wang, P.; Feitosa, F.; Shepherd, D.T.; Ryman, K.D.; Klimstra, W.B.; et al. Mosquito saliva serine protease enhances dissemination of dengue virus into the mammalian host. J. Virol. 2014, 88, 164–175. [Google Scholar] [CrossRef] [Green Version]
- Cox, J.; Mota, J.; Sukupolvi-Petty, S.; Diamond, M.S.; Rico-Hesse, R. Mosquito bite delivery of dengue virus enhances immunogenicity and pathogenesis in humanized mice. J. Virol. 2012, 86, 7637–7649. [Google Scholar] [CrossRef] [Green Version]
- Moser, L.A.; Lim, P.Y.; Styer, L.M.; Kramer, L.D.; Bernard, K.A. Parameters of Mosquito-Enhanced West Nile Virus Infection. J. Virol. 2016, 90, 292–299. [Google Scholar] [CrossRef] [Green Version]
- Pingen, M.; Bryden, S.R.; Pondeville, E.; Schnettler, E.; Kohl, A.; Merits, A.; Fazakerley, J.K.; Graham, G.J.; McKimmie, C.S. Host Inflammatory Response to Mosquito Bites Enhances the Severity of Arbovirus Infection. Immunity 2016, 44, 1455–1469. [Google Scholar] [CrossRef] [Green Version]
- Schmid, M.A.; Glasner, D.R.; Shah, S.; Michlmayr, D.; Kramer, L.D.; Harris, E. Mosquito Saliva Increases Endothelial Permeability in the Skin, Immune Cell Migration, and Dengue Pathogenesis during Antibody-Dependent Enhancement. PLoS Pathog. 2016, 12, e1005676. [Google Scholar] [CrossRef] [Green Version]
- Uraki, R.; Hastings, A.K.; Marin-Lopez, A.; Sumida, T.; Takahashi, T.; Grover, J.R.; Iwasaki, A.; Hafler, D.A.; Montgomery, R.R.; Fikrig, E. Aedes aegypti AgBR1 antibodies modulate early Zika virus infection of mice. Nat. Microbiol. 2019, 4, 948–955. [Google Scholar] [CrossRef]
- Hastings, A.K.; Uraki, R.; Gaitsch, H.; Dhaliwal, K.; Stanley, S.; Sproch, H.; Williamson, E.; MacNeil, T.; Marin-Lopez, A.; Hwang, J.; et al. Aedes aegypti NeSt1 Protein Enhances Zika Virus Pathogenesis by Activating Neutrophils. J. Virol. 2019, 93, e00395-19. [Google Scholar] [CrossRef] [Green Version]
- Fong, S.W.; Kini, R.M.; Ng, L.F.P. Mosquito Saliva Reshapes Alphavirus Infection and Immunopathogenesis. J. Virol. 2018, 92, e01004-17. [Google Scholar] [CrossRef] [Green Version]
- Lefteri, D.A.; Bryden, S.R.; Pingen, M.; Terry, S.; McCafferty, A.; Beswick, E.F.; Georgiev, G.; Van der Laan, M.; Mastrullo, V.; Campagnolo, P.; et al. Mosquito saliva enhances virus infection through sialokinin-dependent vascular leakage. Proc. Natl. Acad. Sci. USA 2022, 119, e2114309119. [Google Scholar] [CrossRef]
- Schuijt, T.J.; Coumou, J.; Narasimhan, S.; Dai, J.; Deponte, K.; Wouters, D.; Brouwer, M.; Oei, A.; Roelofs, J.J.; van Dam, A.P.; et al. A tick mannose-binding lectin inhibitor interferes with the vertebrate complement cascade to enhance transmission of the lyme disease agent. Cell Host Microbe 2011, 10, 136–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, R.; Oliveira, F. The immune response to sand fly salivary proteins and its influence on leishmania immunity. Front. Immunol. 2012, 3, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, M.B.; Lahon, A.; Arya, R.P.; Kneubehl, A.R.; Spencer Clinton, J.L.; Paust, S.; Rico-Hesse, R. Mosquito saliva alone has profound effects on the human immune system. PLoS Negl. Trop. Dis. 2018, 12, e0006439. [Google Scholar] [CrossRef] [PubMed]
- Surasombatpattana, P.; Ekchariyawat, P.; Hamel, R.; Patramool, S.; Thongrungkiat, S.; Denizot, M.; Delaunay, P.; Thomas, F.; Luplertlop, N.; Yssel, H.; et al. Aedes aegypti saliva contains a prominent 34-kDa protein that strongly enhances dengue virus replication in human keratinocytes. J. Invest. Dermatol. 2014, 134, 281–284. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Guo, X.; Shen, C.; Hao, X.; Sun, P.; Li, P.; Xu, T.; Hu, C.; Rose, O.; Zhou, H.; et al. Salivary factor LTRIN from Aedes aegypti facilitates the transmission of Zika virus by interfering with the lymphotoxin-beta receptor. Nat. Immunol. 2018, 19, 342–353. [Google Scholar] [CrossRef]
- Sun, P.; Nie, K.; Zhu, Y.; Liu, Y.; Wu, P.; Liu, Z.; Du, S.; Fan, H.; Chen, C.H.; Zhang, R.; et al. A mosquito salivary protein promotes flavivirus transmission by activation of autophagy. Nat. Commun. 2020, 11, 260. [Google Scholar] [CrossRef] [Green Version]
- Mallard, F.; Antony, C.; Tenza, D.; Salamero, J.; Goud, B.; Johannes, L. Direct pathway from early/recycling endosomes to the Golgi apparatus revealed through the study of shiga toxin B-fragment transport. J. Cell Biol. 1998, 143, 973–990. [Google Scholar] [CrossRef]
- Wanasen, N.; Nussenzveig, R.H.; Champagne, D.E.; Soong, L.; Higgs, S. Differential modulation of murine host immune response by salivary gland extracts from the mosquitoes Aedes aegypti and Culex quinquefasciatus. Med. Vet. Entomol. 2004, 18, 191–199. [Google Scholar] [CrossRef]
- Schneider, B.S.; Soong, L.; Coffey, L.L.; Stevenson, H.L.; McGee, C.E.; Higgs, S. Aedes aegypti saliva alters leukocyte recruitment and cytokine signaling by antigen-presenting cells during West Nile virus infection. PLoS ONE 2010, 5, e11704. [Google Scholar] [CrossRef]
- Conway, M.J.; Londono-Renteria, B.; Troupin, A.; Watson, A.M.; Klimstra, W.B.; Fikrig, E.; Colpitts, T.M. Aedes aegypti D7 Saliva Protein Inhibits Dengue Virus Infection. PLoS Negl. Trop. Dis. 2016, 10, e0004941. [Google Scholar] [CrossRef] [Green Version]
- Reagan, K.L.; Machain-Williams, C.; Wang, T.; Blair, C.D. Immunization of mice with recombinant mosquito salivary protein D7 enhances mortality from subsequent West Nile virus infection via mosquito bite. PLoS Negl. Trop. Dis. 2012, 6, e1935. [Google Scholar] [CrossRef] [Green Version]
- McCracken, M.K.; Christofferson, R.C.; Grasperge, B.J.; Calvo, E.; Chisenhall, D.M.; Mores, C.N. Aedes aegypti salivary protein “aegyptin” co-inoculation modulates dengue virus infection in the vertebrate host. Virology 2014, 468–470, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Collins, N.D.; Barrett, A.D. Live Attenuated Yellow Fever 17D Vaccine: A Legacy Vaccine Still Controlling Outbreaks In Modern Day. Curr. Infect. Dis. Rep. 2017, 19, 14. [Google Scholar] [CrossRef]
- Yun, S.-I.; Lee, Y.-M. Japanese encephalitis. Hum. Vaccines Immunother. 2014, 10, 263–279. [Google Scholar] [CrossRef] [Green Version]
- Satchidanandam, V. Japanese Encephalitis Vaccines. Curr. Treat. Options Infect. Dis. 2020, 12, 375–386. [Google Scholar] [CrossRef]
- Thomas, S.J.; Yoon, I.K. A review of Dengvaxia(R): Development to deployment. Hum Vaccin Immunother. 2019, 15, 2295–2314. [Google Scholar] [CrossRef] [Green Version]
- Takeda’s QDENGA®▼ (Dengue Tetravalent Vaccine [Live, Attenuated]) Approved for Use in European Union. Takeda. 2022. Available online: https://www.takeda.com/newsroom/newsreleases/2022/takedas-qdenga-dengue-tetravalent-vaccine-live-attenuated-approved-for-use-in-european-union (accessed on 31 December 2022).
- Takeda’s QDENGA®▼ (Dengue Tetravalent Vaccine [Live, Attenuated]) Approved in Indonesia for Use Regardless of Prior Dengue Exposure. Takeda. 2022. Available online: https://www.takeda.com/newsroom/newsreleases/2022/takedas-qdenga-dengue-tetravalent-vaccine-live-attenuated-approved-in-indonesia-for-use-regardless-of-prior-dengue-exposure (accessed on 31 December 2022).
- Thomas, R.E.; Lorenzetti, D.L.; Spragins, W.; Jackson, D.; Williamson, T. Reporting rates of yellow fever vaccine 17D or 17DD-associated serious adverse events in pharmacovigilance data bases: Systematic review. Curr. Drug Saf. 2011, 6, 145–154. [Google Scholar] [CrossRef]
- Barrett, A.D.; Teuwen, D.E. Yellow fever vaccine-how does it work and why do rare cases of serious adverse events take place? Curr. Opin. Immunol. 2009, 21, 308–313. [Google Scholar] [CrossRef]
- Dengue Vaccine Safety Update. Available online: https://www.who.int/groups/global-advisory-committee-on-vaccine-safety/topics/dengue-vaccines/safety-update (accessed on 31 December 2022).
- First FDA-Approved Vaccine for the Prevention of Dengue Disease in Endemic Regions. Available online: https://www.fda.gov/news-events/press-announcements/first-fda-approved-vaccine-prevention-dengue-disease-endemic-regions (accessed on 31 December 2022).
- CDC. ZIKV and WNV Cases in the United States. Available online: https://www.cdc.gov/westnile/index.html (accessed on 31 December 2022).
- Diamond, M.S.; Ledgerwood, J.E.; Pierson, T.C. Zika Virus Vaccine Development: Progress in the Face of New Challenges. Annu. Rev. Med. 2019, 70, 121–135. [Google Scholar] [CrossRef]
- Shan, C.; Xie, X.; Shi, P.Y. Zika Virus Vaccine: Progress and Challenges. Cell Host Microbe 2018, 24, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Scherwitzl, I.; Mongkolsapaja, J.; Screaton, G. Recent advances in human flavivirus vaccines. Curr. Opin. Virol. 2017, 23, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Zhu, B.; Fu, Z.F.; Chen, H.; Cao, S. Immune evasion strategies of flaviviruses. Vaccine 2013, 31, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.G.; Alvarez, M.; Halstead, S.B. Secondary infection as a risk factor for dengue hemorrhagic fever/dengue shock syndrome: An historical perspective and role of antibody-dependent enhancement of infection. Arch. Virol. 2013, 158, 1445–1459. [Google Scholar] [CrossRef] [PubMed]
- Garg, H.; Yeh, R.; Watts, D.M.; Mehmetoglu-Gurbuz, T.; Resendes, R.; Parsons, B.; Gonzales, F.; Joshi, A. Enhancement of Zika virus infection by antibodies from West Nile virus seropositive individuals with no history of clinical infection. BMC Immunol. 2021, 22, 5. [Google Scholar] [CrossRef] [PubMed]
- Bardina, S.V.; Bunduc, P.; Tripathi, S.; Duehr, J.; Frere, J.J.; Brown, J.A.; Nachbagauer, R.; Foster, G.A.; Krysztof, D.; Tortorella, D.; et al. Enhancement of Zika virus pathogenesis by preexisting antiflavivirus immunity. Science 2017, 356, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Machain-Williams, C.; Reagan, K.; Wang, T.; Zeidner, N.S.; Blair, C.D. Immunization with Culex tarsalis mosquito salivary gland extract modulates West Nile virus infection and disease in mice. Viral. Immunol. 2013, 26, 84–92. [Google Scholar] [CrossRef] [Green Version]
- Marin-Lopez, A.; Wang, Y.; Jiang, J.; Ledizet, M.; Fikrig, E. AgBR1 and NeSt1 antisera protect mice from Aedes aegypti-borne Zika infection. Vaccine 2021, 39, 1675–1679. [Google Scholar] [CrossRef]
- Wang, Y.; Marin-Lopez, A.; Jiang, J.; Ledizet, M.; Fikrig, E. Vaccination with Aedes aegypti AgBR1 Delays Lethal Mosquito-Borne Zika Virus Infection in Mice. Vaccines 2020, 8, 145. [Google Scholar] [CrossRef] [Green Version]
- Uraki, R.; Hastings, A.K.; Brackney, D.E.; Armstrong, P.M.; Fikrig, E. AgBR1 antibodies delay lethal Aedes aegypti-borne West Nile virus infection in mice. NPJ Vaccines 2019, 4, 23. [Google Scholar] [CrossRef] [Green Version]
- Londono-Renteria, B.; Troupin, A.; Colpitts, T.M. Arbovirosis and potential transmission blocking vaccines. Parasit Vectors 2016, 9, 516. [Google Scholar] [CrossRef] [Green Version]
- Troupin, A.; Grippin, C.; Colpitts, T.M. Flavivirus Pathogenesis in the Mosquito Transmission Vector. Curr. Clin. Microbiol. Rep. 2017, 4, 115–123. [Google Scholar] [CrossRef]
- Londono-Renteria, B.; Troupin, A.; Conway, M.J.; Vesely, D.; Ledizet, M.; Roundy, C.M.; Cloherty, E.; Jameson, S.; Vanlandingham, D.; Higgs, S.; et al. Dengue Virus Infection of Aedes aegypti Requires a Putative Cysteine Rich Venom Protein. PLoS Pathog. 2015, 11, e1005202. [Google Scholar] [CrossRef] [Green Version]
- Sri-In, C.; Weng, S.C.; Chen, W.Y.; Wu-Hsieh, B.A.; Tu, W.C.; Shiao, S.H. A salivary protein of Aedes aegypti promotes dengue-2 virus replication and transmission. Insect. Biochem. Mol Biol. 2019, 111, 103181. [Google Scholar] [CrossRef]
- Manning, J.E.; Oliveira, F.; Coutinho-Abreu, I.V.; Herbert, S.; Meneses, C.; Kamhawi, S.; Baus, H.A.; Han, A.; Czajkowski, L.; Rosas, L.A.; et al. Safety and immunogenicity of a mosquito saliva peptide-based vaccine: A randomised, placebo-controlled, double-blind, phase 1 trial. Lancet 2020, 395, 1998–2007. [Google Scholar] [CrossRef]
- Friedman-Klabanoff, D.J.; Birkhold, M.; Short, M.T.; Wilson, T.R.; Meneses, C.R.; Lacsina, J.R.; Oliveira, F.; Kamhawi, S.; Valenzuela, J.G.; Hunsberger, S.; et al. Safety and immunogenicity of AGS-v PLUS, a mosquito saliva peptide vaccine against arboviral diseases: A randomized, double-blind, placebo-controlled Phase 1 trial. EBioMedicine 2022, 86, 104375. [Google Scholar] [CrossRef]
- National Center for Emerging and Zoonotic Infectious Diseases (NCEZID), Division of Vector-Borne Diseases (DVBD): Centers for Disease Control and Prevention. 2022. Available online: https://www.cdc.gov/ncezid/dvbd/index.html (accessed on 2 November 2022).
- Sagna, A.B.; Yobo, M.C.; Elanga Ndille, E.; Remoue, F. New Immuno-Epidemiological Biomarker of Human Exposure to Aedes Vector Bites: From Concept to Applications. Trop. Med. Infect. Dis. 2018, 3, 80. [Google Scholar] [CrossRef] [Green Version]
- Wasinpiyamongkol, L.; Patramool, S.; Luplertlop, N.; Surasombatpattana, P.; Doucoure, S.; Mouchet, F.; Seveno, M.; Remoue, F.; Demettre, E.; Brizard, J.P.; et al. Blood-feeding and immunogenic Aedes aegypti saliva proteins. Proteomics 2010, 10, 1906–1916. [Google Scholar] [CrossRef]
- Doucoure, S.; Mouchet, F.; Cournil, A.; Le Goff, G.; Cornelie, S.; Roca, Y.; Giraldez, M.G.; Simon, Z.B.; Loayza, R.; Misse, D.; et al. Human antibody response to Aedes aegypti saliva in an urban population in Bolivia: A new biomarker of exposure to Dengue vector bites. Am. J. Trop. Med. Hyg. 2012, 87, 504–510. [Google Scholar] [CrossRef] [Green Version]
- Machain-Williams, C.; Mammen, M.P.; Zeidner, N.S., Jr.; Beaty, B.J.; Prenni, J.E.; Nisalak, A.; Blair, C.D. Association of human immune response to Aedes aegypti salivary proteins with dengue disease severity. Parasite Immunol. 2012, 34, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Manning, J.E.; Chea, S.; Parker, D.M.; Bohl, J.A.; Lay, S.; Mateja, A.; Man, S.; Nhek, S.; Ponce, A.; Sreng, S.; et al. Development of Inapparent Dengue Associated With Increased Antibody Levels to Aedes aegypti Salivary Proteins: A Longitudinal Dengue Cohort in Cambodia. J. Infect. Dis. 2022, 226, 1327–1337. [Google Scholar] [CrossRef]
- Buezo Montero, S.; Gabrieli, P.; Severini, F.; Picci, L.; Di Luca, M.; Forneris, F.; Facchinelli, L.; Ponzi, M.; Lombardo, F.; Arca, B. Analysis in a murine model points to IgG responses against the 34k2 salivary proteins from Aedes albopictus and Aedes aegypti as novel promising candidate markers of host exposure to Aedes mosquitoes. PLoS Negl. Trop. Dis. 2019, 13, e0007806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fustec, B.; Phanitchat, T.; Aromseree, S.; Pientong, C.; Thaewnongiew, K.; Ekalaksananan, T.; Cerqueira, D.; Poinsignon, A.; Elguero, E.; Bangs, M.J.; et al. Serological biomarker for assessing human exposure to Aedes mosquito bites during a randomized vector control intervention trial in northeastern Thailand. PLoS Negl. Trop. Dis. 2021, 15, e0009440. [Google Scholar] [CrossRef] [PubMed]
- Buezo Montero, S.; Gabrieli, P.; Poinsignon, A.; Zamble, B.Z.H.; Lombardo, F.; Remoue, F.; Arca, B. Human IgG responses to the Aedes albopictus 34k2 salivary protein: Analyses in Reunion Island and Bolivia confirm its suitability as marker of host exposure to the tiger mosquito. Parasit Vectors 2022, 15, 260. [Google Scholar] [CrossRef] [PubMed]
- Olajiga, O.M.; Marin-Lopez, A.; Cardenas, J.C.; Gutierrez-Silva, L.Y.; Gonzales-Pabon, M.U.; Maldonado-Ruiz, L.P.; Worges, M.; Fikrig, E.; Park, Y.; Londono-Renteria, B. Aedes aegypti anti-salivary proteins IgG levels in a cohort of DENV-like symptoms subjects from a dengue-endemic region in Colombia. Front. Epidemiol. 2022, 2. [Google Scholar] [CrossRef]
- Londono-Renteria, B.L.; Shakeri, H.; Rozo-Lopez, P.; Conway, M.J.; Duggan, N.; Jaberi-Douraki, M.; Colpitts, T.M. Serosurvey of Human Antibodies Recognizing Aedes aegypti D7 Salivary Proteins in Colombia. Front. Public Health 2018, 6, 111. [Google Scholar] [CrossRef] [Green Version]
- Brosseau, L.; Drame, P.M.; Besnard, P.; Toto, J.C.; Foumane, V.; Le Mire, J.; Mouchet, F.; Remoue, F.; Allan, R.; Fortes, F.; et al. Human antibody response to Anopheles saliva for comparing the efficacy of three malaria vector control methods in Balombo, Angola. PLoS ONE 2012, 7, e44189. [Google Scholar] [CrossRef] [Green Version]
- Noukpo, M.H.; Damien, G.B.; Elanga-N’Dille, E.; Sagna, A.B.; Drame, P.M.; Chaffa, E.; Boussari, O.; Corbel, V.; Akogbeto, M.; Remoue, F. Operational Assessment of Long-Lasting Insecticidal Nets by Using an Anopheles Salivary Biomarker of Human-Vector Contact. Am. J. Trop. Med. Hyg. 2016, 95, 1376–1382. [Google Scholar] [CrossRef] [Green Version]
- Pollard, E.J.M.; Patterson, C.; Russell, T.L.; Apairamo, A.; Oscar, J.; Arca, B.; Drakeley, C.; Burkot, T.R. Human exposure to Anopheles farauti bites in the Solomon Islands is not associated with IgG antibody response to the gSG6 salivary protein of Anopheles gambiae. Malar. J. 2019, 18, 334. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, C.; Lombardo, F.; Ronca, R.; Mangano, V.; Sirima, S.B.; Nebie, I.; Fiorentino, G.; Modiano, D.; Arca, B. Differential antibody response to the Anopheles gambiae gSG6 and cE5 salivary proteins in individuals naturally exposed to bites of malaria vectors. Parasit Vectors 2014, 7, 549. [Google Scholar] [CrossRef]
- Ali, Z.M.; Bakli, M.; Fontaine, A.; Bakkali, N.; Vu Hai, V.; Audebert, S.; Boublik, Y.; Pages, F.; Remoue, F.; Rogier, C.; et al. Assessment of Anopheles salivary antigens as individual exposure biomarkers to species-specific malaria vector bites. Malar. J. 2012, 11, 439. [Google Scholar] [CrossRef] [Green Version]
- Londono-Renteria, B.; Drame, P.M.; Montiel, J.; Vasquez, A.M.; Tobon-Castano, A.; Taylor, M.; Vizcaino, L.; Lenhart, A.A.E. Identification and Pilot Evaluation of Salivary Peptides from Anopheles albimanus as Biomarkers for Bite Exposure and Malaria Infection in Colombia. Int. J. Mol. Sci. 2020, 21, 691. [Google Scholar] [CrossRef] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marín-López, A.; Raduwan, H.; Chen, T.-Y.; Utrilla-Trigo, S.; Wolfhard, D.P.; Fikrig, E. Mosquito Salivary Proteins and Arbovirus Infection: From Viral Enhancers to Potential Targets for Vaccines. Pathogens 2023, 12, 371. https://doi.org/10.3390/pathogens12030371
Marín-López A, Raduwan H, Chen T-Y, Utrilla-Trigo S, Wolfhard DP, Fikrig E. Mosquito Salivary Proteins and Arbovirus Infection: From Viral Enhancers to Potential Targets for Vaccines. Pathogens. 2023; 12(3):371. https://doi.org/10.3390/pathogens12030371
Chicago/Turabian StyleMarín-López, Alejandro, Hamidah Raduwan, Tse-Yu Chen, Sergio Utrilla-Trigo, David P. Wolfhard, and Erol Fikrig. 2023. "Mosquito Salivary Proteins and Arbovirus Infection: From Viral Enhancers to Potential Targets for Vaccines" Pathogens 12, no. 3: 371. https://doi.org/10.3390/pathogens12030371