
Carvacrol as a Stimulant of the Expression of Key Genes of the Ginsenoside Biosynthesis Pathway and Its Effect on the Production of Ginseng Saponins in Panax quinquefolium Hairy Root Cultures
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Effect of Carvacrol Treatment on Expression Level of Studied Genes in Hairy Roots of P. quinquefolium
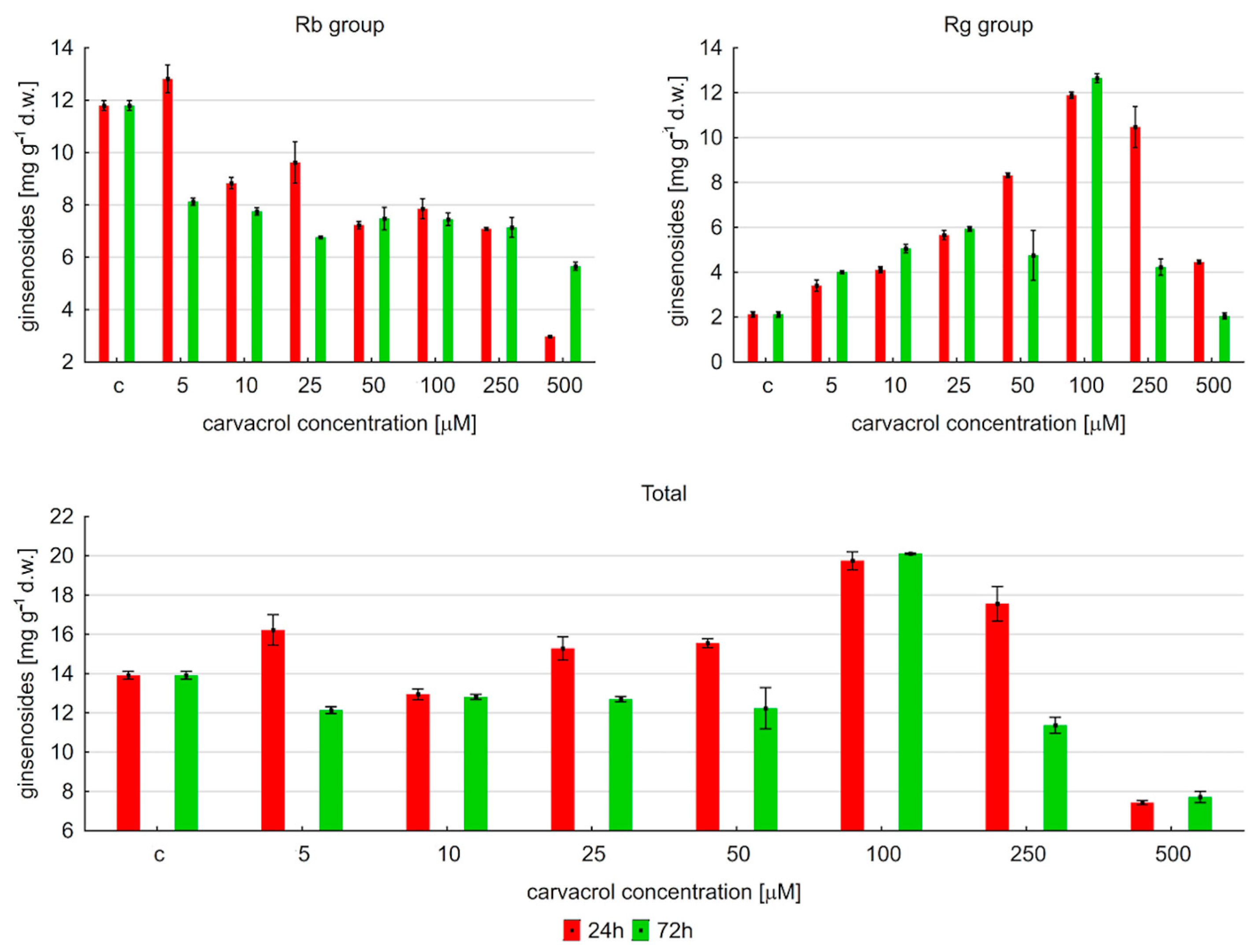
2.2. Effect of Carvacrol Treatment on Ginsenoside Production in Hairy Roots of P. quinquefolium
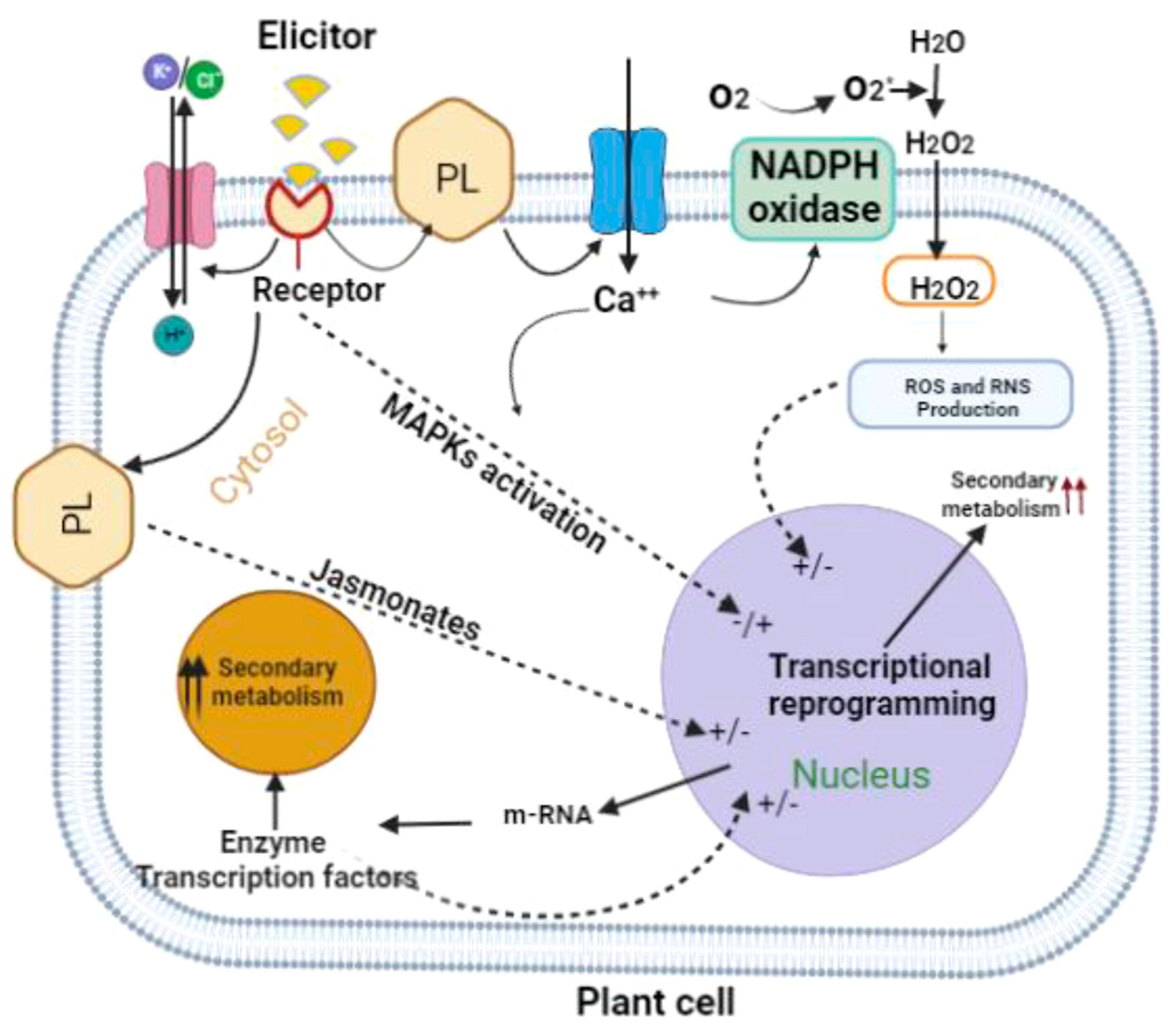
3. Discussion
4. Materials and Methods
4.1. Hairy Roots of P. quinquefolium Cultured in Shaken Flasks
4.2. Elicitation of Hairy Root Cultures of P. quinquefolium
4.3. RNA Isolation and Reverse Transcriptase (RT)-PCR Analysis
4.4. Qualitative Analysis
4.5. Quantitative Analysis by Real-Time PCR
4.6. Determination of Ginsenoside Content
4.6.1. Sample Preparation
4.6.2. Quantitative Analysis of Ginsenosides with the HPLC Method
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Divekar, P.A.; Narayana, S.; Divekar, B.A.; Kumar, R.; Gadratagi, B.G.; Ray, A.; Singh, A.K.; Rani, V.; Singh, V.; Singh, A.K.; et al. Plant Secondary Metabolites as Defense Tools against Herbivores for Sustainable Crop Protection. Int. J. Mol. Sci. 2022, 23, 2690. [Google Scholar] [CrossRef] [PubMed]
- Yeshi, K.; Crayn, D.; Ritmejerytė, E.; Wangchuk, P. Plant Secondary Metabolites Produced in Response to Abiotic Stresses Has Potential Application in Pharmaceutical Product Development. Molecules 2022, 27, 313. [Google Scholar] [CrossRef] [PubMed]
- Laftouhi, A.; Eloutassi, N.; Ech-Chihbi, E.; Rais, Z.; Abdellaoui, A.; Taleb, A.; Beniken, M.; Nafidi, H.-A.; Salamatullah, A.M.; Bourhia, M.; et al. The Impact of Environmental Stress on the Secondary Metabolites and the Chemical Compositions of the Essential Oils from Some Medicinal Plants Used as Food Supplements. Sustainability 2023, 15, 7842. [Google Scholar] [CrossRef]
- Hussein, R.A.; El-Anssary, A.A. Plants Secondary Metabolites: The Key Drivers of the Pharmacological Actions of Medicinal Plants. In Herbal Medicine; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Lee, S.; Rhee, D.-K. Effects of Ginseng on Stress-Related Depression, Anxiety, and the Hypothalamic–Pituitary–Adrenal Axis. J. Ginseng Res. 2017, 41, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Verstraeten, S.L.; Lorent, J.H.; Mingeot-Leclercq, M.-P. Lipid Membranes as Key Targets for the Pharmacological Actions of Ginsenosides. Front. Pharmacol. 2020, 11, 576887. [Google Scholar] [CrossRef]
- Shi, Z.-Y.; Zeng, J.-Z.; Wong, A.S.T. Chemical Structures and Pharmacological Profiles of Ginseng Saponins. Molecules 2019, 24, 2443. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Wu, H.; Zhang, H.; Zhao, Q.; Yin, X.; Zheng, D.; Li, C.; Kim, M.; Kim, P.; Xue, Z.; et al. Highly Efficient Production of Diverse Rare Ginsenosides Using Combinatorial Biotechnology. Biotechnol. Bioeng. 2020, 117, 1615–1627. [Google Scholar] [CrossRef]
- Piao, X.M.; Huo, Y.; Kang, J.P.; Mathiyalagan, R.; Zhang, H.; Yang, D.U.; Kim, M.; Yang, D.C.; Kang, S.C.; Wang, Y.P. Diversity of Ginsenoside Profiles Produced by Various Processing Technologies. Molecules 2020, 25, 4390. [Google Scholar] [CrossRef]
- Gupta, R.; Jain, D. Transforming plant secondary metabolite production through the application of PTC and elicitor technology. J. Adv. Sci. Res. 2022, 13, 75–86. [Google Scholar] [CrossRef]
- Humbal, A.; Pathak, B. Influence of Exogenous Elicitors on the Production of Secondary Metabolite in Plants: A Review (“VSI: Secondary Metabolites”). Plant Stress 2023, 8, 100166. [Google Scholar] [CrossRef]
- Ramirez-Estrada, K.; Vidal-Limon, H.; Hidalgo, D.; Moyano, E.; Golenioswki, M.; Cusidó, R.; Palazon, J. Elicitation, an Effective Strategy for the Biotechnological Production of Bioactive High-Added Value Compounds in Plant Cell Factories. Molecules 2016, 21, 182. [Google Scholar] [CrossRef]
- Kairuz, E.; Pérez-Alonso, N.; Capote-Pérez, A.; Pérez-Pérez, A.; Espinosa-Antón, A.A.; Angenon, G.; Jiménez, E.; Chong-Pérez, B. Enhancement of Cardenolide Production in Transgenic Digitalis purpurea L. by Expressing a Progesterone-5β-Reductase from Arabidopsis thaliana L. Ind. Crops Prod. 2020, 146, 112166. [Google Scholar] [CrossRef]
- Mohanlall, V. Plant Cell Culture Systems for the Production of Secondary Metabolites—A Review. IOSR J. Biotechnol. Biochem. 2020, 6, 35–47. [Google Scholar]
- Halder, M.; Sarkar, S.; Jha, S. Elicitation: A Biotechnological Tool for Enhanced Production of Secondary Metabolites in Hairy Root Cultures. Eng. Life Sci. 2019, 19, 880–895. [Google Scholar] [CrossRef]
- Asl, K.R.; Hosseini, B.; Sharafi, A.; Palazon, J. Influence of Nano-Zinc Oxide on Tropane Alkaloid Production, H6h Gene Transcription and Antioxidant Enzyme Activity in Hyoscyamus reticulatus L. Hairy Roots. Eng. Life Sci. 2019, 19, 73–89. [Google Scholar] [CrossRef]
- Hanafy, M.S.; Matter, M.A.; Asker, M.S.; Rady, M.R. Production of Indole Alkaloids in Hairy Root Cultures of Catharanthus roseus L. and Their Antimicrobial Activity. S. Afr. J. Bot. 2016, 105, 9–18. [Google Scholar] [CrossRef]
- Kim, O.T.; Bang, K.H.; Kim, Y.C.; Hyun, D.Y.; Kim, M.Y.; Cha, S.W. Upregulation of Ginsenoside and Gene Expression Related to Triterpene Biosynthesis in Ginseng Hairy Root Cultures Elicited by Methyl Jasmonate. Plant Cell Tissue Organ Cult. PCTOC 2009, 98, 25–33. [Google Scholar] [CrossRef]
- Palazón, J.; Cusidó, R.M.; Bonfill, M.; Mallol, A.; Moyano, E.; Morales, C.; Piñol, M.T. Elicitation of Different Panax ginseng Transformed Root Phenotypes for an Improved Ginsenoside Production. Plant Physiol. Biochem. 2003, 41, 1019–1025. [Google Scholar] [CrossRef]
- Sivakumar, G.; Yu, K.; Hahn, E. Optimization of Organic Nutrients for Ginseng Hairy Roots Production in Large-Scale Bioreactors. Curr. Sci. 2005, 89, 641–649. [Google Scholar]
- Yu, K.-W.; Gao, W.-Y.; Son, S.-H.; Paek, K.-Y. Improvement of Ginsenoside Production by Jasmonic Acid and Some Other Elicitors in Hairy Root Culture of Ginseng (Panax ginseng C. A. Meyer). Vitro Cell. Dev. Biol.-Plant 2000, 36, 424–428. [Google Scholar] [CrossRef]
- Yu, K.-W.; Murthy, H.N.; Hahn, E.-J.; Paek, K.-Y. Ginsenoside Production by Hairy Root Cultures of Panax ginseng: Influence of Temperature and Light Quality. Biochem. Eng. J. 2005, 23, 53–56. [Google Scholar] [CrossRef]
- Liang, Y.; Wu, J.; Li, Y.; Li, J.; Ouyang, Y.; He, Z.; Zhao, S. Enhancement of Ginsenoside Biosynthesis and Secretion by Tween 80 in Panax ginseng Hairy Roots. Biotechnol. Appl. Biochem. 2015, 62, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Han, J.Y.; Jo, H.-J.; Choi, Y.E. Overexpression of the Squalene Epoxidase Gene (PgSE1) Resulted in Enhanced Production of Ginsenosides and Phytosterols in Transgenic Ginseng. Plant Biotechnol. Rep. 2020, 14, 673–682. [Google Scholar] [CrossRef]
- Han, J.-Y.; In, J.-G.; Kwon, Y.-S.; Choi, Y.-E. Regulation of Ginsenoside and Phytosterol Biosynthesis by RNA Interferences of Squalene Epoxidase Gene in Panax ginseng. Phytochemistry 2010, 71, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-K.; Kim, Y.B.; Uddin, M.R.; Lee, S.; Kim, S.-U.; Park, S.U. Enhanced Triterpene Accumulation in Panax ginseng Hairy Roots Overexpressing Mevalonate-5-Pyrophosphate Decarboxylase and Farnesyl Pyrophosphate Synthase. ACS Synth. Biol. 2014, 3, 773–779. [Google Scholar] [CrossRef]
- Lee, M.-H.; Jeong, J.-H.; Seo, J.-W.; Shin, C.-G.; Kim, Y.-S.; In, J.-G.; Yang, D.-C.; Yi, J.-S.; Choi, Y.-E. Enhanced Triterpene and Phytosterol Biosynthesis in Panax ginseng Overexpressing Squalene Synthase Gene. Plant Cell Physiol. 2004, 45, 976–984. [Google Scholar] [CrossRef]
- Kochan, E.; Królicka, A.; Chmiel, A. Growth and Ginsenoside Production in Panax Quinquefolium Hairy Roots Cultivated in Flasks and Nutrient Sprinkle Bioreactor. Acta Physiol. Plant. 2012, 34, 1513–1518. [Google Scholar] [CrossRef]
- Kochan, E.; Szymczyk, P.; Kuźma, Ł.; Lipert, A.; Szymańska, G. Yeast Extract Stimulates Ginsenoside Production in Hairy Root Cultures of American Ginseng Cultivated in Shake Flasks and Nutrient Sprinkle Bioreactors. Molecules 2017, 22, 880. [Google Scholar] [CrossRef]
- Kochan, E.; Balcerczak, E.; Lipert, A.; Szymańska, G.; Szymczyk, P. Methyl Jasmonate as a Control Factor of the Synthase Squalene Gene Promoter and Ginsenoside Production in American Ginseng Hairy Root Cultured in Shake Flasks and a Nutrient Sprinkle Bioreactor. Ind. Crops Prod. 2018, 115, 182–193. [Google Scholar] [CrossRef]
- Kochan, E.; Balcerczak, E.; Szymczyk, P.; Sienkiewicz, M.; Zielińska-Bliźniewska, H.; Szymańska, G. Abscisic Acid Regulates the 3-Hydroxy-3-Methylglutaryl CoA Reductase Gene Promoter and Ginsenoside Production in Panax Quinquefolium Hairy Root Cultures. Int. J. Mol. Sci. 2019, 20, 1310. [Google Scholar] [CrossRef]
- Osaili, T.M.; Dhanasekaran, D.K.; Zeb, F.; Faris, M.E.; Naja, F.; Radwan, H.; Cheikh Ismail, L.; Hasan, H.; Hashim, M.; Obaid, R.S. A Status Review on Health-Promoting Properties and Global Regulation of Essential Oils. Molecules 2023, 28, 1809. [Google Scholar] [CrossRef] [PubMed]
- Kochan, E.; Szymańska, G.; Kwiatkowski, P.; Sienkiewicz, M. Linalool as a Novel Natural Factor Enhancing Ginsenoside Production in Hairy Root Cultures of American Ginseng. Plant Cell Tissue Organ Cult. PCTOC 2023, 153, 285–293. [Google Scholar] [CrossRef]
- Kochan, E.; Szymczyk, P.; Kuźma, Ł.; Szymańska, G.; Wajs-Bonikowska, A.; Bonikowski, R.; Sienkiewicz, M. The Increase of Triterpene Saponin Production Induced by Trans-Anethole in Hairy Root Cultures of Panax Quinquefolium. Molecules 2018, 23, 2674. [Google Scholar] [CrossRef]
- Matias, E.F.F.; Pereira, A.P.D.; Braz, A.V.D.O.; Rodrigues, M.C.; Silva, J.D.L.; Maia, P.A.A.; Santos, S.C.D.; Rebelo, R.A.; Begnini, I.M.; Silva, L.E.D.; et al. Chemical Profile and Biological Activities of Essential Oil from Piper Arboreum for Development and Improvement of Mouthwash. Molecules 2022, 27, 6408. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Aslam, M.; Alsagaby, S.A.; Saeed, F.; Ahmad, I.; Afzaal, M.; Arshad, M.U.; Abdelgawad, M.A.; El-Ghorab, A.H.; Khames, A.; et al. Therapeutic Application of Carvacrol: A Comprehensive Review. Food Sci. Nutr. 2022, 10, 3544–3561. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowski, P.; Sienkiewicz, M.; Pruss, A.; Łopusiewicz, Ł.; Arszyńska, N.; Wojciechowska-Koszko, I.; Kilanowicz, A.; Kot, B.; Dołęgowska, B. Antibacterial and Anti-Biofilm Activities of Essential Oil Compounds against New Delhi Metallo-β-Lactamase-1-Producing Uropathogenic Klebsiella Pneumoniae Strains. Antibiotics 2022, 11, 147. [Google Scholar] [CrossRef]
- Langhansova, L. Increased Ginsenosides Production by Elicitation of In Vitro Cultivated Panax ginseng Adventitious Roots. Med. Aromat. Plants 2014, 3, 2167-0412. [Google Scholar] [CrossRef]
- Paek, K.-Y.; Murthy, H.N.; Hahn, E.-J.; Zhong, J.-J. Large Scale Culture of Ginseng Adventitious Roots for Production of Ginsenosides. In Biotechnology in China I: From Bioreaction to Bioseparation and Bioremediation; Zhong, J.-J., Bai, F.-W., Zhang, W., Eds.; Advances in Biochemical Engineering/Biotechnology; Springer: Berlin/Heidelberg, Germany, 2009; pp. 151–176. ISBN 978-3-540-88415-6. [Google Scholar]
- Yang, J.-L.; Hu, Z.-F.; Zhang, T.-T.; Gu, A.-D.; Gong, T.; Zhu, P. Progress on the Studies of the Key Enzymes of Ginsenoside Biosynthesis. Molecules 2018, 23, 589. [Google Scholar] [CrossRef]
- Kim, O.T.; Bang, K.H.; Jung, S.J.; Kim, Y.C.; Hyun, D.Y.; Kim, S.H.; Cha, S.W. Molecular Characterization of Ginseng Farnesyl Diphosphate Synthase Gene and Its Up-Regulation by Methyl Jasmonate. Biol. Plant. 2010, 54, 47–53. [Google Scholar] [CrossRef]
- Li, X.; Liu, J.; Zuo, T.; Hu, Y.; Li, Z.; Wang, H.; Xu, X.; Yang, W.; Guo, D. Advances and Challenges in Ginseng Research from 2011 to 2020: The Phytochemistry, Quality Control, Metabolism, and Biosynthesis. Nat. Prod. Rep. 2022, 39, 875–909. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Yeung, E.C.; Hahn, E.-J.; Paek, K.-Y. Combined Effects of Phytohormone, Indole-3-Butyric Acid, and Methyl Jasmonate on Root Growth and Ginsenoside Production in Adventitious Root Cultures of Panax ginseng C.A. Meyer. Biotechnol. Lett. 2007, 29, 1789–1792. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gao, W.; Zuo, B.; Zhang, L.; Huang, L. Effect of Methyl Jasmonate on the Ginsenoside Content of Panax ginseng Adventitious Root Cultures and on the Genes Involved in Triterpene Biosynthesis. Res. Chem. Intermed. 2013, 39, 1973–1980. [Google Scholar] [CrossRef]
- Yao, L.; Wang, J.; Sun, J.; He, J.; Paek, K.; Park, S.-Y.; Huang, L.; Gao, W. A WRKY Transcription Factor, PgWRKY4X, Positively Regulates Ginsenoside Biosynthesis by Activating Squalene Epoxidase Transcription in Panax ginseng. Ind. Crops Prod. 2020, 154, 112671. [Google Scholar] [CrossRef]
- Han, J.Y.; Kwon, Y.S.; Yang, D.C.; Jung, Y.R.; Choi, Y.E. Expression and RNA Interference-Induced Silencing of the Dammarenediol Synthase Gene in Panax ginseng. Plant Cell Physiol. 2006, 47, 1653–1662. [Google Scholar] [CrossRef]
- Liang, Y.; Zhao, S.; Zhang, X. Antisense Suppression of Cycloartenol Synthase Results in Elevated Ginsenoside Levels in Panax ginseng Hairy Roots. Plant Mol. Biol. Rep. 2009, 27, 298–304. [Google Scholar] [CrossRef]
- Mohanan, P.; Subramaniyam, S.; Mathiyalagan, R.; Yang, D.-C. Molecular Signaling of Ginsenosides Rb1, Rg1, and Rg3 and Their Mode of Actions. J. Ginseng Res. 2018, 42, 123–132. [Google Scholar] [CrossRef]
- Han, J.-Y.; Kim, H.-J.; Kwon, Y.-S.; Choi, Y.-E. The Cyt P450 Enzyme CYP716A47 Catalyzes the Formation of Protopanaxadiol from Dammarenediol-II during Ginsenoside Biosynthesis in Panax ginseng. Plant Cell Physiol. 2011, 52, 2062–2073. [Google Scholar] [CrossRef] [PubMed]
- Shin, B.-K.; Kwon, S.W.; Park, J.H. Chemical Diversity of Ginseng Saponins from Panax ginseng. J. Ginseng Res. 2015, 39, 287–298. [Google Scholar] [CrossRef]
- Park, S.-B.; Chun, J.-H.; Ban, Y.-W.; Han, J.Y.; Choi, Y.E. Alteration of Panax ginseng Saponin Composition by Overexpression and RNA Interference of the Protopanaxadiol 6-Hydroxylase Gene (CYP716A53v2). J. Ginseng Res. 2016, 40, 47–54. [Google Scholar] [CrossRef]
- Hou, M.; Wang, R.; Zhao, S.; Wang, Z. Ginsenosides in Panax Genus and Their Biosynthesis. Acta Pharm. Sin. B 2021, 11, 1813–1834. [Google Scholar] [CrossRef]
- Chen, S.; Luo, H.; Li, Y.; Sun, Y.; Wu, Q.; Niu, Y.; Song, J.; Lv, A.; Zhu, Y.; Sun, C.; et al. 454 EST Analysis Detects Genes Putatively Involved in Ginsenoside Biosynthesis in Panax ginseng. Plant Cell Rep. 2011, 30, 1593–1601. [Google Scholar] [CrossRef]
- Jung, S.-C.; Kim, W.; Park, S.C.; Jeong, J.; Park, M.K.; Lim, S.; Lee, Y.; Im, W.-T.; Lee, J.H.; Choi, G.; et al. Two Ginseng UDP-Glycosyltransferases Synthesize Ginsenoside Rg3 and Rd. Plant Cell Physiol. 2014, 55, 2177–2188. [Google Scholar] [CrossRef]
- Kang, K.B.; Jayakodi, M.; Lee, Y.S.; Nguyen, V.B.; Park, H.-S.; Koo, H.J.; Choi, I.Y.; Kim, D.H.; Chung, Y.J.; Ryu, B.; et al. Identification of Candidate UDP-Glycosyltransferases Involved in Protopanaxadiol-Type Ginsenoside Biosynthesis in Panax ginseng. Sci. Rep. 2018, 8, 11744. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhu, Y.; Guo, X.; Sun, C.; Luo, H.; Song, J.; Li, Y.; Wang, L.; Qian, J.; Chen, S. Transcriptome Analysis Reveals Ginsenosides Biosynthetic Genes, microRNAs and Simple Sequence Repeats in Panax ginseng C. A. Meyer. BMC Genom. 2013, 14, 245. [Google Scholar] [CrossRef]
- Luo, H.; Sun, C.; Sun, Y.; Wu, Q.; Li, Y.; Song, J.; Niu, Y.; Cheng, X.; Xu, H.; Li, C.; et al. Analysis of the Transcriptome of Panax Notoginseng Root Uncovers Putative Triterpene Saponin-Biosynthetic Genes and Genetic Markers. BMC Genom. 2011, 12, S5. [Google Scholar] [CrossRef]
- Sun, C.; Li, Y.; Wu, Q.; Luo, H.; Sun, Y.; Song, J.; Lui, E.M.; Chen, S. De Novo Sequencing and Analysis of the American Ginseng Root Transcriptome Using a GS FLX Titanium Platform to Discover Putative Genes Involved in Ginsenoside Biosynthesis. BMC Genom. 2010, 11, 262. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, S.; Kim, J.; Mijakovic, I.; Jung, K.-H.; Choi, G.; Kim, S.-C.; Kim, Y.-J. Triterpenoid-Biosynthetic UDP-Glycosyltransferases from Plants. Biotechnol. Adv. 2019, 37, 107394. [Google Scholar] [CrossRef] [PubMed]
- Gamborg, O.L.; Miller, R.A.; Ojima, K. Nutrient Requirements of Suspension Cultures of Soybean Root Cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- Kochan, E.; Szymczyk, P.; Kuźma, Ł.; Szymańska, G. Nitrogen and Phosphorus as the Factors Affecting Ginsenoside Production in Hairy Root Cultures of Panax Quinquefolium Cultivated in Shake Flasks and Nutrient Sprinkle Bioreactor. Acta Physiol. Plant. 2016, 38, 149. [Google Scholar] [CrossRef]
- Kibbe, W.A. OligoCalc: An Online Oligonucleotide Properties Calculator. Nucleic Acids Res. 2007, 35, W43–W46. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Zakrajsek, B.A. Effect of Experimental Treatment on Housekeeping Gene Expression: Validation by Real-Time, Quantitative RT-PCR. J. Biochem. Biophys. Methods 2000, 46, 69–81. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ginsenosides | Elicitation 24 h | Elicitation 72 h |
|---|---|---|
| Rg1 | −0.25 | −0.06 |
| Re | 0.72 | 0.04 |
| Rg2 | 0.65 | 0.47 |
| Rb1 | −0.91 | −0.57 |
| Rc | −0.83 | −0.59 |
| Rb2 | −0.81 | −0.69 |
| Rb3 | −0.82 | −0.61 |
| Rd | −0.90 | −0.87 |
| Rg group | 0.79 | 0.08 |
| Rb group | −0.87 | −0.74 |
| Total | 0.09 | −0.40 |
| Gene | Primers | GenBank Accession Number |
|---|---|---|
| DS | Forward: 5′CACAGCTGAAGCGCTAAAGT3′ Reverse: 5′ATATGGTTTTGGAACTGGAGGCT3′ | KC316048.1 |
| FPS | Forward: 5′ATAGATTTGATCACCACCCTT G3′ Reverse: 5′AGTATGTTTCTCCAGATCTTCGC3′ | GQ401664.1 |
| SS | Forward: 5′ATGCTGAAGTCCAAGGTTGAC3′ Reverse: 5′ATTGTATCCTGACTCATTCTCG3′ | AM 182456.1 |
| actin | Forward: 5′AAGCCCAATCGAAGAGAGGTA3′ Reverse: 5′ATCTTCTCCCTGTTGGCCTT3′ | KF 699319.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kochan, E.; Sienkiewicz, M.; Szmajda-Krygier, D.; Balcerczak, E.; Szymańska, G. Carvacrol as a Stimulant of the Expression of Key Genes of the Ginsenoside Biosynthesis Pathway and Its Effect on the Production of Ginseng Saponins in Panax quinquefolium Hairy Root Cultures. Int. J. Mol. Sci. 2024, 25, 909. https://doi.org/10.3390/ijms25020909
Kochan E, Sienkiewicz M, Szmajda-Krygier D, Balcerczak E, Szymańska G. Carvacrol as a Stimulant of the Expression of Key Genes of the Ginsenoside Biosynthesis Pathway and Its Effect on the Production of Ginseng Saponins in Panax quinquefolium Hairy Root Cultures. International Journal of Molecular Sciences. 2024; 25(2):909. https://doi.org/10.3390/ijms25020909
Chicago/Turabian StyleKochan, Ewa, Monika Sienkiewicz, Dagmara Szmajda-Krygier, Ewa Balcerczak, and Grażyna Szymańska. 2024. "Carvacrol as a Stimulant of the Expression of Key Genes of the Ginsenoside Biosynthesis Pathway and Its Effect on the Production of Ginseng Saponins in Panax quinquefolium Hairy Root Cultures" International Journal of Molecular Sciences 25, no. 2: 909. https://doi.org/10.3390/ijms25020909