
The Effect of DREADD Activation of Leptin Receptor Positive Neurons in the Nucleus of the Solitary Tract on Sleep Disordered Breathing

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
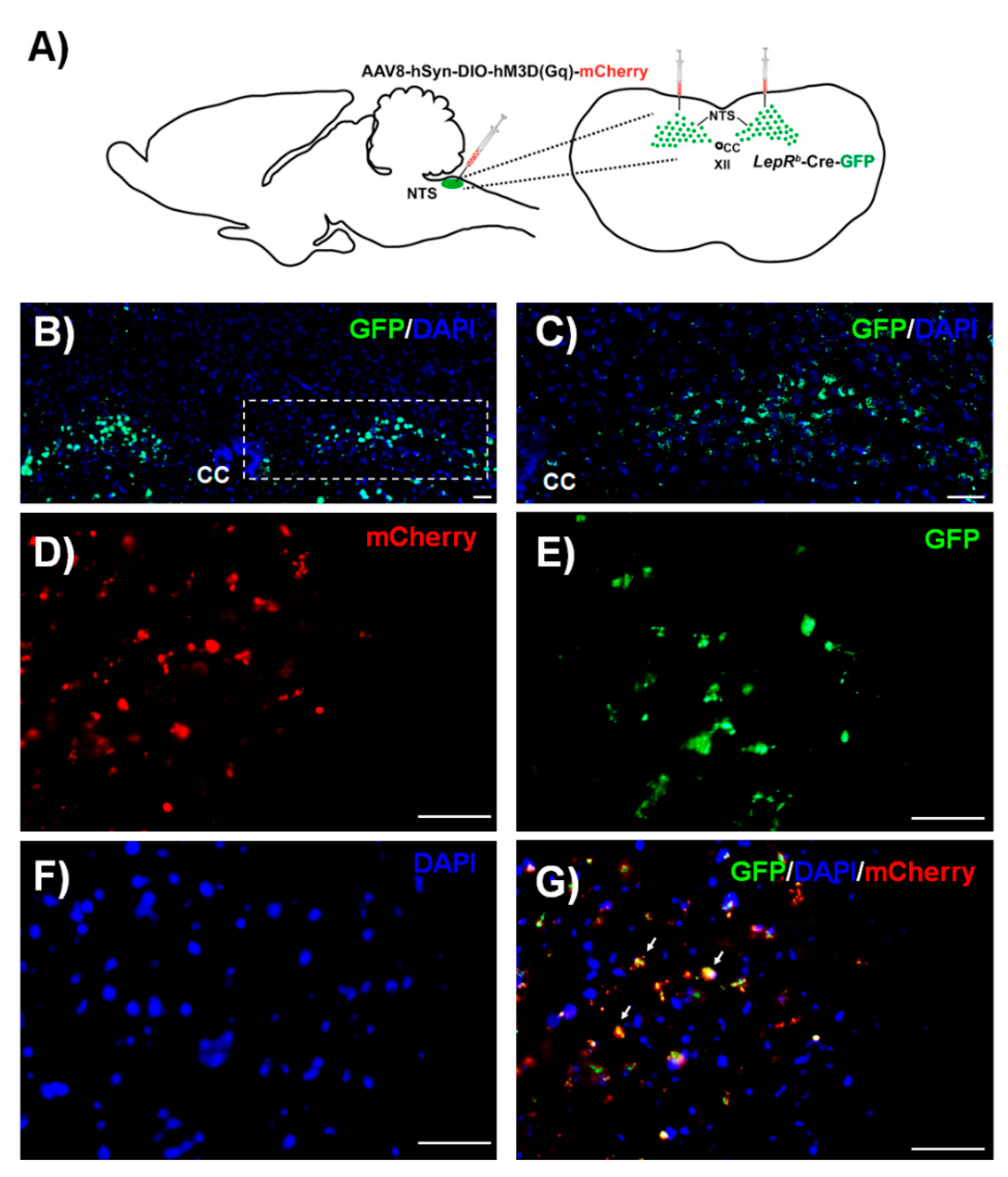
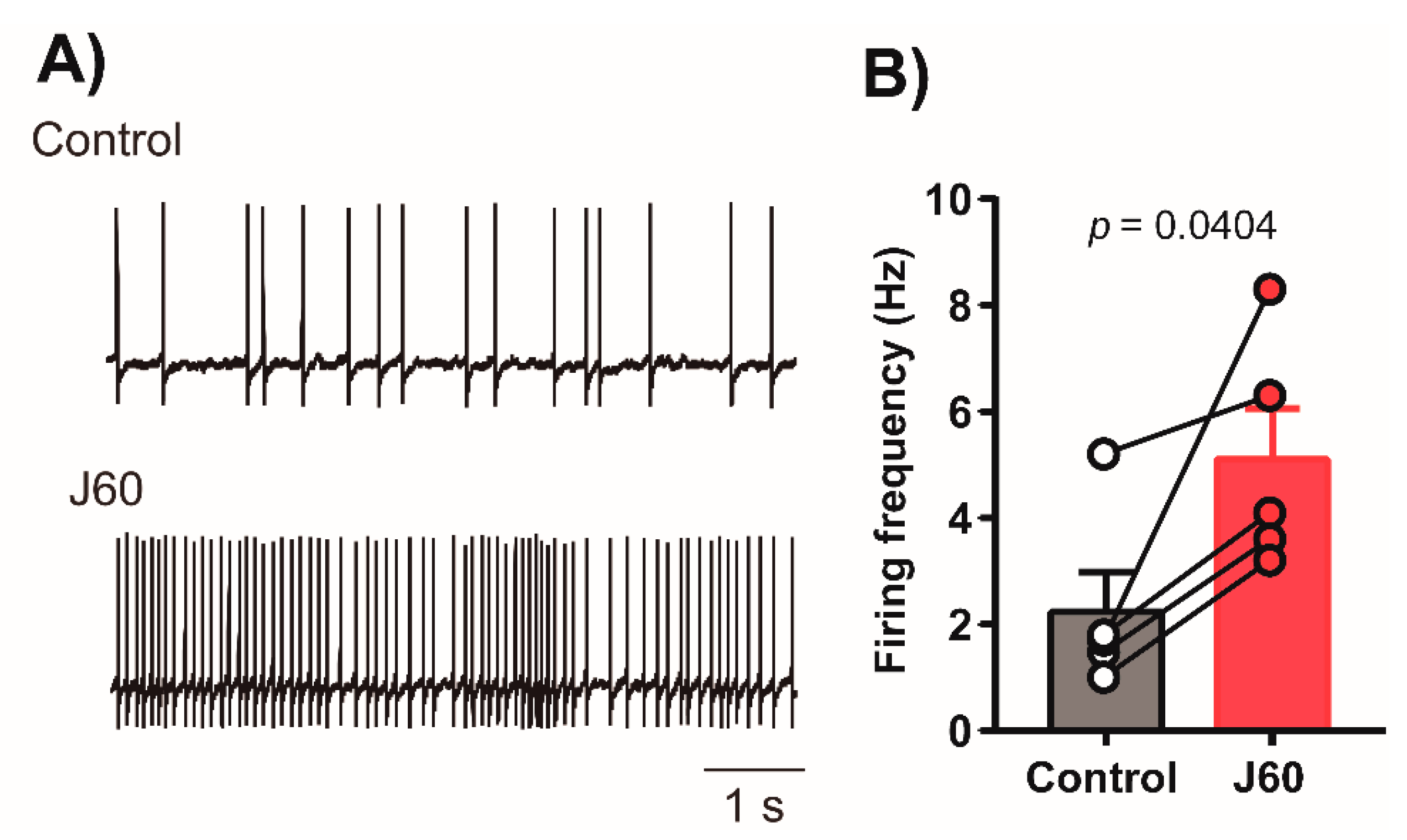
2.1. Histology and Electrophysiology
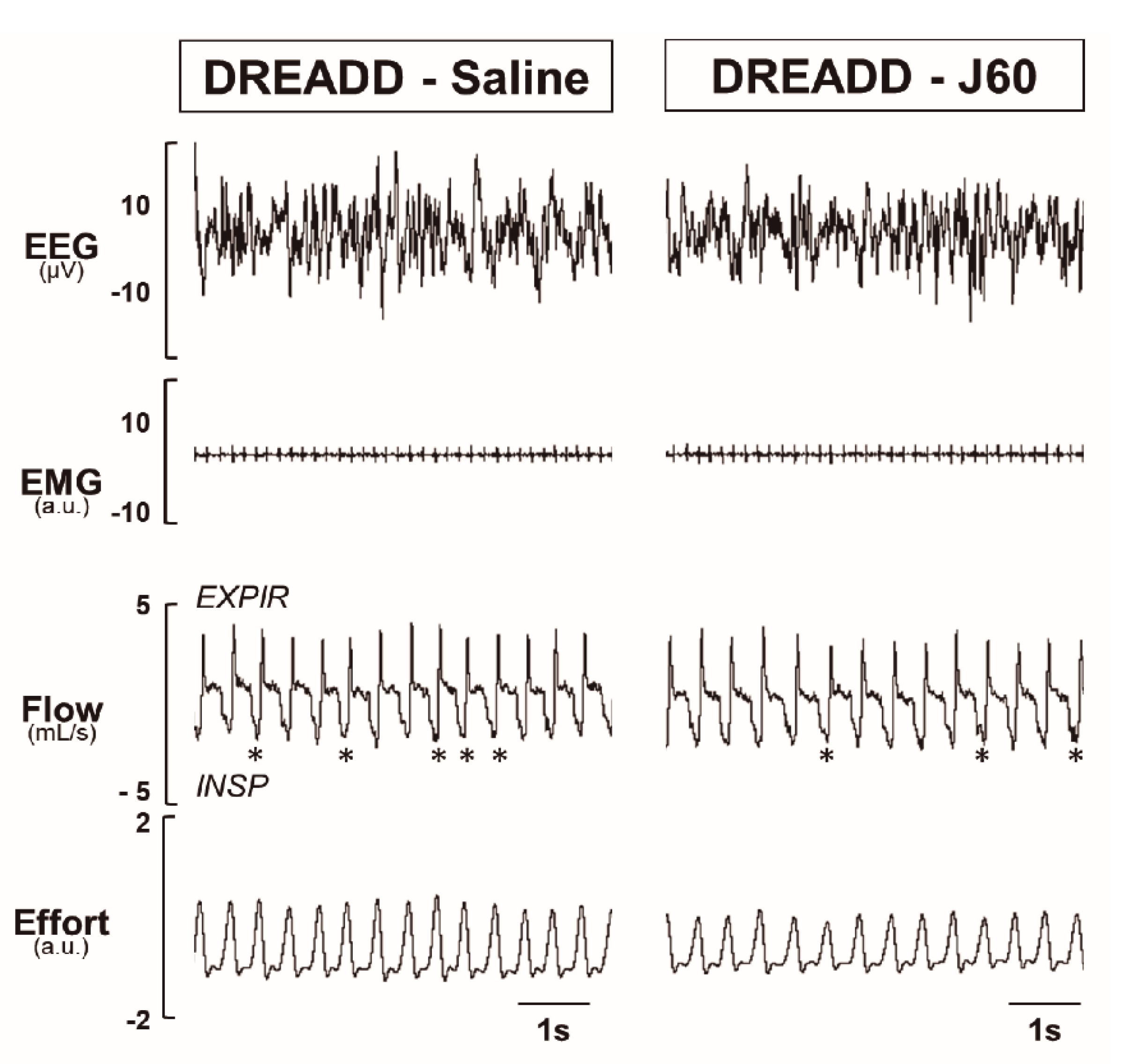
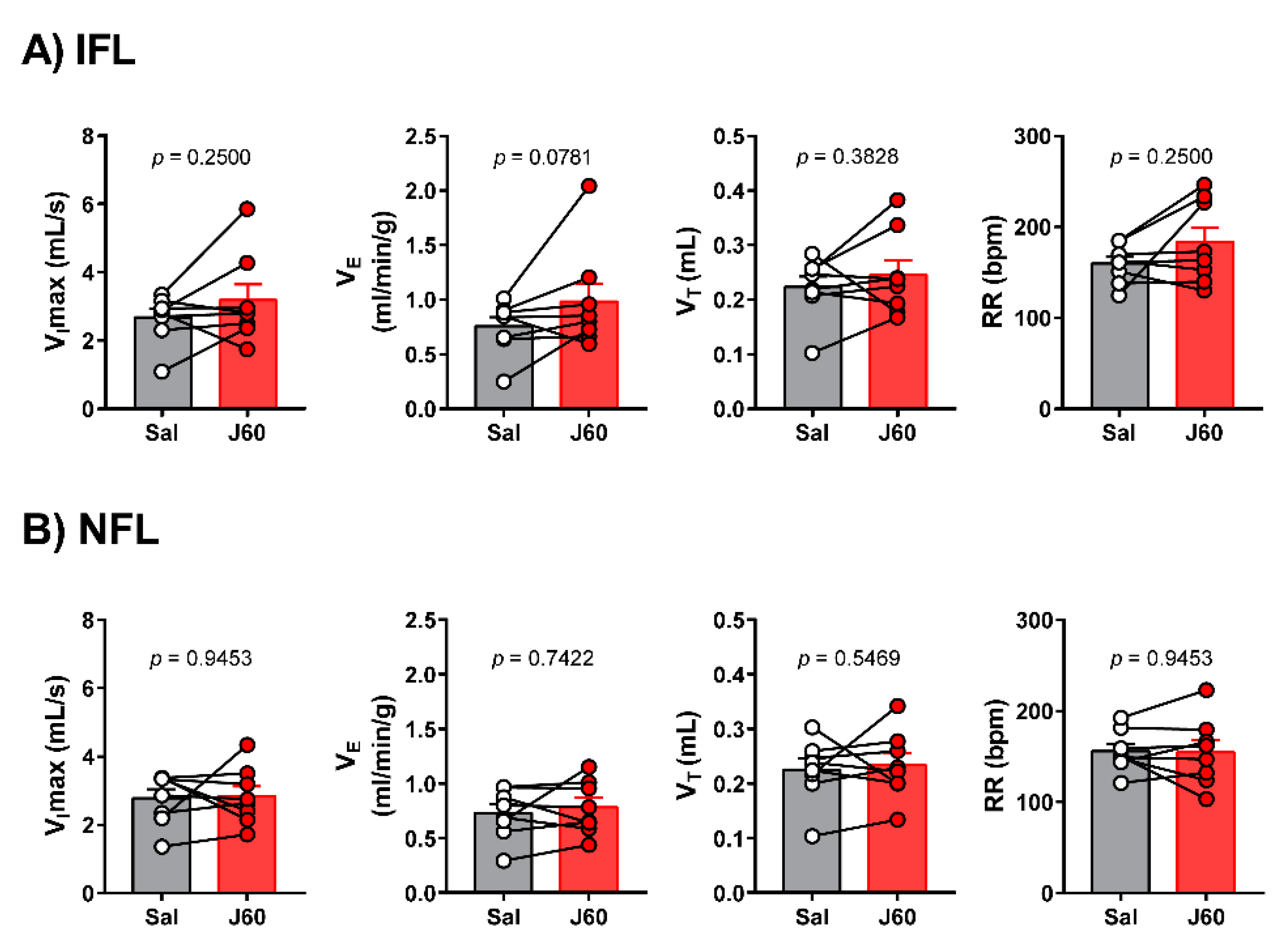
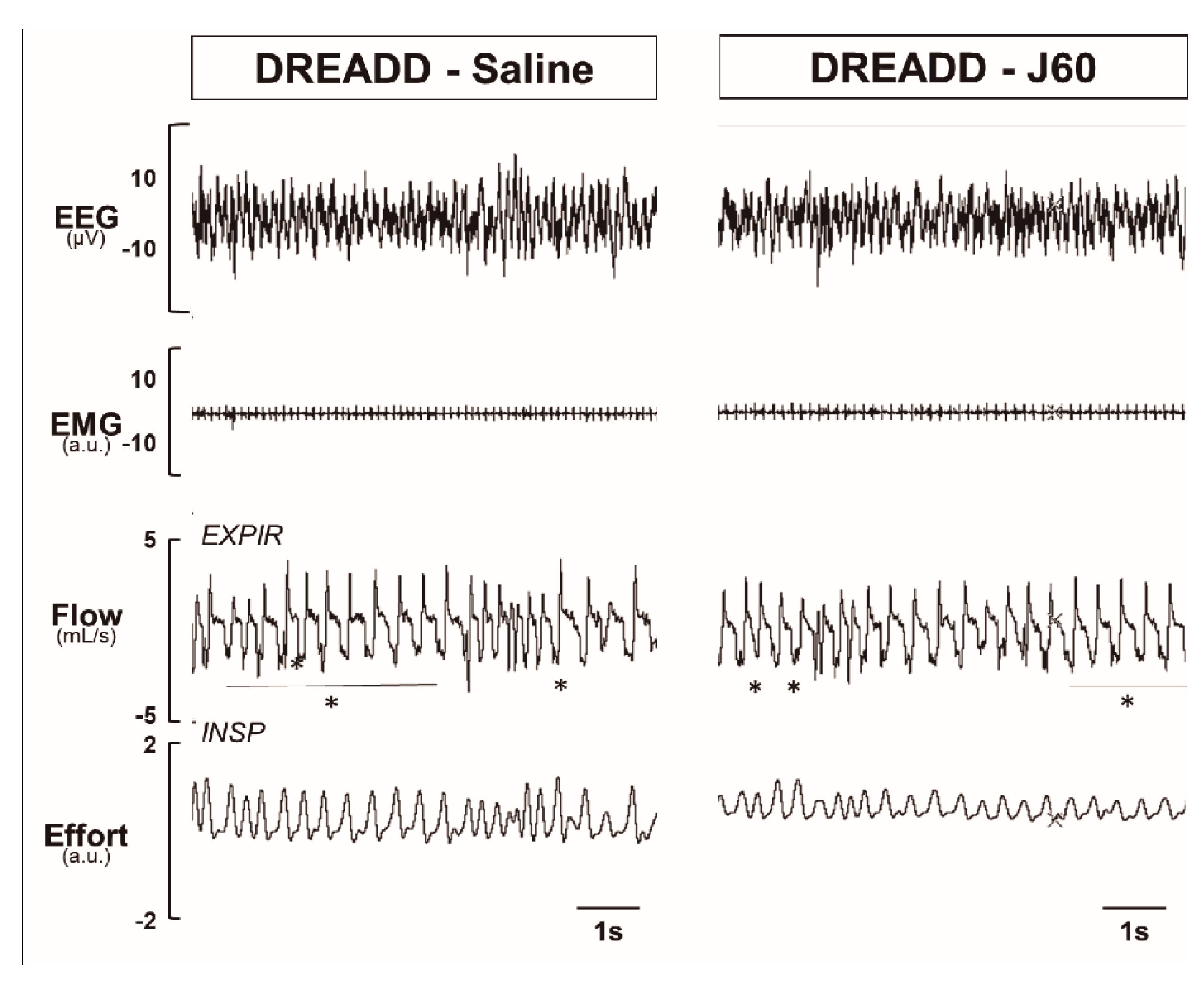
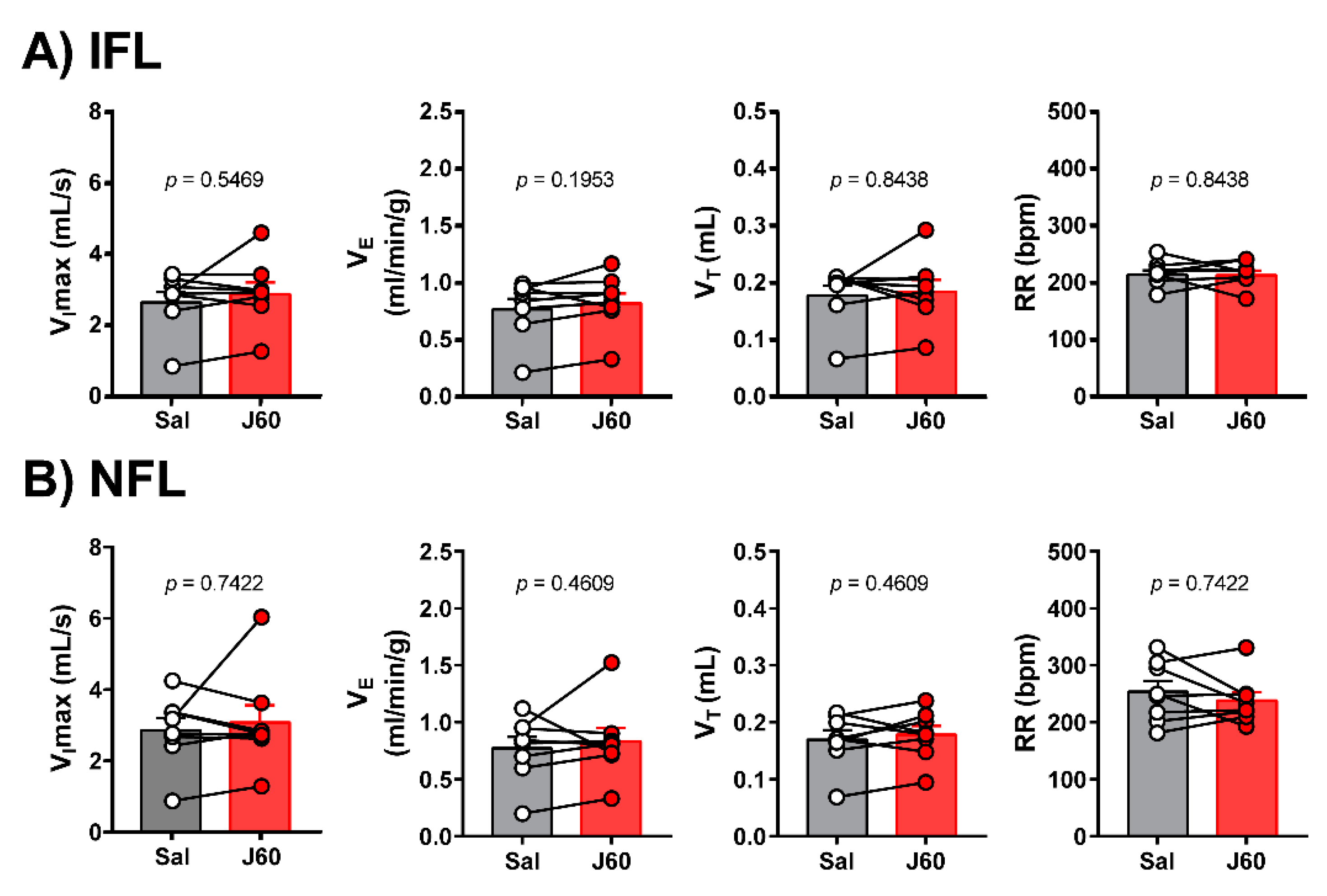
2.2. Sleep Studies
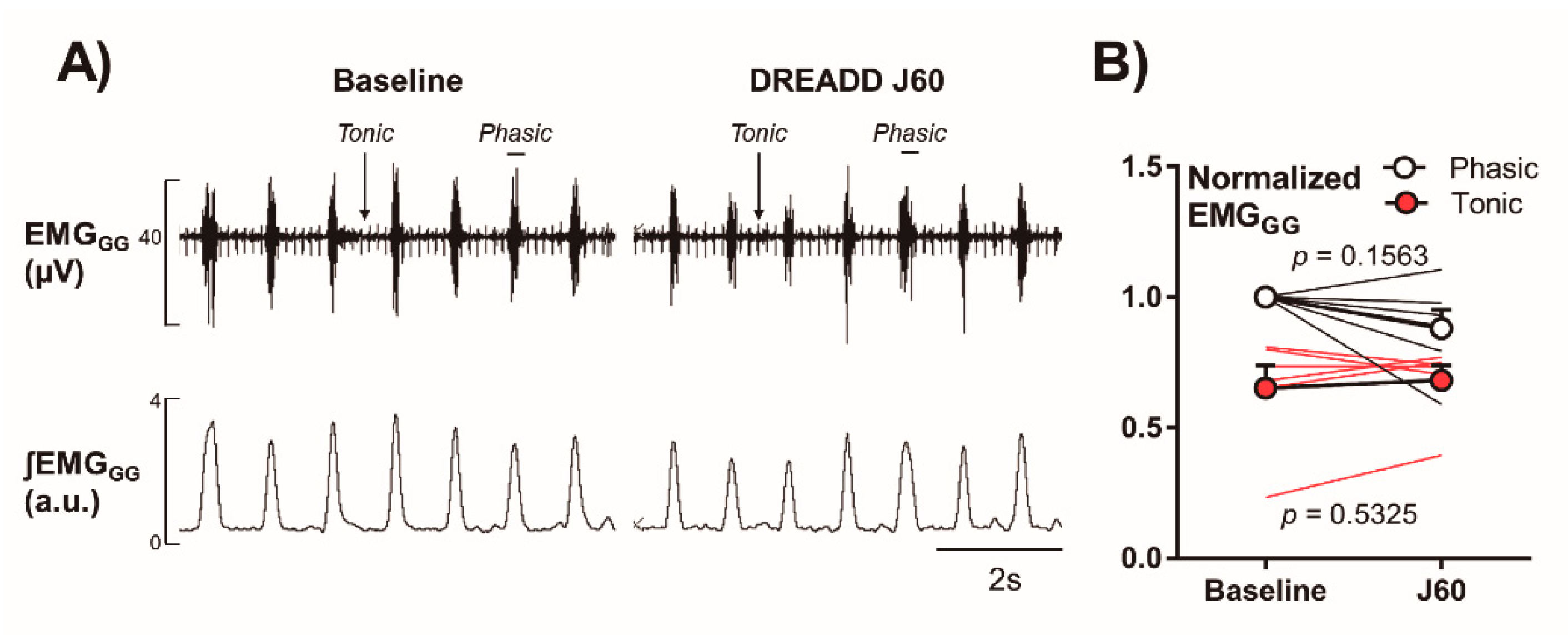
2.3. Electromyography of the Genioglossus Muscle (EMGGG)
3. Discussion
3.1. The Effect of Activation of LEPRb Positive NTS Neurons on Breathing during Sleep
3.2. The Effect of Activation of LEPRb Positive NTS Neurons in the Upper Airway Pattency
4. Materials and Methods
4.1. Animals and Ethical Approval
4.2. Viral Vector Administration
4.3. Sleep Studies
4.4. Electromyography of the Genioglossus Muscle (EMGGG)
4.5. Selective Expression and Activation of DREADD in NTS LEPRb Positive Neurons
4.6. Immunofluorescence
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peppard, P.E.; Young, T.; Barnet, J.H.; Palta, M.; Hagen, E.W.; Hla, K.M. Increased Prevalence of Sleep-Disordered Breathing in Adults. Am. J. Epidemiol. 2013, 177, 1006–1014. [Google Scholar] [CrossRef] [Green Version]
- Heinzer, R.; Vat, S.; Marques-Vidal, P.; Marti-Soler, H.; Andries, D.; Tobback, N.; Mooser, V.; Preisig, M.; Malhotra, A.; Waeber, G.; et al. Prevalence of Sleep-Disordered Breathing in the General Population: The HypnoLaus Study. Lancet Respir. Med. 2015, 3, 310–318. [Google Scholar] [CrossRef] [Green Version]
- Young, T.; Palta, M.; Dempsey, J.; Skatrud, J.; Weber, S.; Badr, S. The Occurrence of Sleep-Disordered Breathing among Middle-Aged Adults. N. Engl. J. Med. 1993, 328, 1230–1235. [Google Scholar] [CrossRef] [Green Version]
- Tufik, S.; Santos-Silva, R.; Taddei, J.A.; Bittencourt, L.R.A. Obstructive Sleep Apnea Syndrome in the Sao Paulo Epidemiologic Sleep Study. Sleep Med. 2010, 11, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Punjabi, N.M.; Sorkin, J.D.; Katzel, L.I.; Goldberg, A.P.; Schwartz, A.R.; Smith, P.L. Sleep-Disordered Breathing and Insulin Resistance in Middle-Aged and Overweight Men. Am. J. Respir. Crit. Care Med. 2002, 165, 677–682. [Google Scholar] [CrossRef]
- Young, T.; Peppard, P.E.; Gottlieb, D.J. Epidemiology of Obstructive Sleep Apnea: A Population Health Perspective. Am. J. Respir. Crit. Care Med. 2002, 165, 1217–1239. [Google Scholar] [CrossRef]
- Mesarwi, O.A.; Sharma, E.V.; Jun, J.C.; Polotsky, V.Y. Metabolic Dysfunction in Obstructive Sleep Apnea: A Critical Examination of Underlying Mechanisms. Sleep Biol. Rhythms 2015, 13, 2–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gami, A.S.; Howard, D.E.; Olson, E.J.; Somers, V.K. Day-Night Pattern of Sudden Death in Obstructive Sleep Apnea. N. Engl. J. Med. 2005, 352, 1206–1214. [Google Scholar] [CrossRef] [PubMed]
- Yaggi, H.K.; Concato, J.; Kernan, W.N.; Lichtman, J.H.; Brass, L.M.; Mohsenin, V. Obstructive Sleep Apnea as a Risk Factor for Stroke and Death. N. Engl. J. Med. 2005, 353, 2034–2041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javaheri, S.; Barbe, F.; Campos-Rodriguez, F.; Dempsey, J.A.; Khayat, R.; Javaheri, S.; Malhotra, A.; Martinez-Garcia, M.A.; Mehra, R.; Pack, A.I.; et al. Sleep Apnea: Types, Mechanisms, and Clinical Cardiovascular Consequences. J. Am. Coll. Cardiol. 2017, 69, 841–858. [Google Scholar] [CrossRef]
- Hirotsu, C.; Haba-Rubio, J.; Togeiro, S.M.; Marques-Vidal, P.; Drager, L.F.; Vollenweider, P.; Waeber, G.; Bittencourt, L.; Tufik, S.; Heinzer, R. Obstructive Sleep Apnoea as a Risk Factor for Incident Metabolic Syndrome: A Joined Episono and HypnoLaus Prospective Cohorts Study. Eur. Respir. J. 2018, 52. [Google Scholar] [CrossRef] [Green Version]
- Gabryelska, A.; Karuga, F.F.; Szmyd, B.; Białasiewicz, P. HIF-1α as a Mediator of Insulin Resistance, T2DM, and Its Complications: Potential Links with Obstructive Sleep Apnea. Front. Physiol. 2020, 11, 1035. [Google Scholar] [CrossRef] [PubMed]
- Kribbs, N.B.; Pack, A.I.; Kline, L.R.; Smith, P.L.; Schwartz, A.R.; Schubert, N.M.; Redline, S.; Henry, J.N.; Getsy, J.E.; Dinges, D.F. Objective Measurement of Patterns of Nasal CPAP Use by Patients with Obstructive Sleep Apnea. Am. Rev. Respir. Dis. 1993, 147, 887–895. [Google Scholar] [CrossRef]
- Yao, Q.; Pho, H.; Kirkness, J.; Ladenheim, E.E.; Bi, S.; Moran, T.H.; Fuller, D.D.; Schwartz, A.R.; Polotsky, V.Y. Localizing Effects of Leptin on Upper Airway and Respiratory Control during Sleep. Sleep 2016, 39, 1097–1106. [Google Scholar] [CrossRef] [PubMed]
- Fleury Curado, T.; Fishbein, K.; Pho, H.; Brennick, M.; Dergacheva, O.; Sennes, L.U.; Pham, L.V.; Ladenheim, E.E.; Spencer, R.; Mendelowitz, D.; et al. Chemogenetic Stimulation of the Hypoglossal Neurons Improves Upper Airway Patency. Sci. Rep. 2017, 7, 44392. [Google Scholar] [CrossRef] [Green Version]
- Fleury Curado, T.; Pho, H.; Berger, S.; Caballero-Eraso, C.; Shin, M.-K.; Sennes, L.U.; Pham, L.; Schwartz, A.R.; Polotsky, V.Y. Sleep-Disordered Breathing in C57BL/6J Mice with Diet-Induced Obesity. Sleep 2018, 41. [Google Scholar] [CrossRef]
- Berger, S.; Pho, H.; Fleury-Curado, T.; Bevans-Fonti, S.; Younas, H.; Shin, M.-K.; Jun, J.C.; Anokye-Danso, F.; Ahima, R.S.; Enquist, L.W.; et al. Intranasal Leptin Relieves Sleep-Disordered Breathing in Mice with Diet-Induced Obesity. Am. J. Respir. Crit. Care Med. 2019, 199, 773–783. [Google Scholar] [CrossRef]
- Kim, L.J.; Freire, C.; Fleury Curado, T.; Jun, J.C.; Polotsky, V.Y. The Role of Animal Models in Developing Pharmacotherapy for Obstructive Sleep Apnea. J. Clin. Med. 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Pho, H.; Berger, S.; Freire, C.; Kim, L.J.; Shin, M.-K.; Streeter, S.R.; Hosamane, N.; Cabassa, M.E.; Anokye-Danso, F.; Dergacheva, O.; et al. Leptin Receptor Expression in the Dorsomedial Hypothalamus Stimulates Breathing during NREM Sleep in Db/Db Mice. Sleep 2021. [Google Scholar] [CrossRef]
- Szmyd, B.; Rogut, M.; Białasiewicz, P.; Gabryelska, A. The Impact of Glucocorticoids and Statins on Sleep Quality. Sleep Med. Rev. 2021, 55, 101380. [Google Scholar] [CrossRef]
- Friedman, J.M. Leptin at 14 y of Age: An Ongoing Story. Am. J. Clin. Nutr. 2009, 89, 973S–979S. [Google Scholar] [CrossRef] [Green Version]
- Halaas, J.L.; Gajiwala, K.S.; Maffei, M.; Cohen, S.L.; Chait, B.T.; Rabinowitz, D.; Lallone, R.L.; Burley, S.K.; Friedman, J.M. Weight-Reducing Effects of the Plasma Protein Encoded by the Obese Gene. Science 1995, 269, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Spiegelman, B.M.; Flier, J.S. Obesity and the Regulation of Energy Balance. Cell 2001, 104, 531–543. [Google Scholar] [CrossRef] [Green Version]
- O’donnell, C.P.; Schaub, C.D.; Haines, A.S.; Berkowitz, D.E.; Tankersley, C.G.; Schwartz, A.R.; Smith, P.L. Leptin Prevents Respiratory Depression in Obesity. Am. J. Respir. Crit. Care Med. 1999, 159, 1477–1484. [Google Scholar] [CrossRef] [PubMed]
- Wauman, J.; Tavernier, J. Leptin Receptor Signaling: Pathways to Leptin Resistance. Front. Biosci. Landmark Ed. 2011, 16, 2771–2793. [Google Scholar] [CrossRef] [Green Version]
- Scott, M.M.; Lachey, J.L.; Sternson, S.M.; Lee, C.E.; Elias, C.F.; Friedman, J.M.; Elmquist, J.K. Leptin Targets in the Mouse Brain. J. Comp. Neurol. 2009, 514, 518–532. [Google Scholar] [CrossRef] [Green Version]
- Pho, H.; Hernandez, A.B.; Arias, R.S.; Leitner, E.B.; Van Kooten, S.; Kirkness, J.P.; Schneider, H.; Smith, P.L.; Polotsky, V.Y.; Schwartz, A.R. The Effect of Leptin Replacement on Sleep-Disordered Breathing in the Leptin-Deficient Ob/Ob Mouse. J. Appl. Physiol. 2016, 120, 78–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Considine, R.V.; Sinha, M.K.; Heiman, M.L.; Kriauciunas, A.; Stephens, T.W.; Nyce, M.R.; Ohannesian, J.P.; Marco, C.C.; McKee, L.J.; Bauer, T.L. Serum Immunoreactive-Leptin Concentrations in Normal-Weight and Obese Humans. N. Engl. J. Med. 1996, 334, 292–295. [Google Scholar] [CrossRef]
- Maffei, M.; Halaas, J.; Ravussin, E.; Pratley, R.E.; Lee, G.H.; Zhang, Y.; Fei, H.; Kim, S.; Lallone, R.; Ranganathan, S. Leptin Levels in Human and Rodent: Measurement of Plasma Leptin and Ob RNA in Obese and Weight-Reduced Subjects. Nat. Med. 1995, 1, 1155–1161. [Google Scholar] [CrossRef]
- Inyushkin, A.N.; Inyushkina, E.M.; Merkulova, N.A. Respiratory Responses to Microinjections of Leptin into the Solitary Tract Nucleus. Neurosci. Behav. Physiol. 2009, 39, 231–240. [Google Scholar] [CrossRef]
- Inyushkina, E.M.; Merkulova, N.A.; Inyushkin, A.N. Mechanisms of the Respiratory Activity of Leptin at the Level of the Solitary Tract Nucleus. Neurosci. Behav. Physiol. 2010, 40, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Do, J.; Chang, Z.; Sekerková, G.; McCrimmon, D.R.; Martina, M. A Leptin-Mediated Neural Mechanism Linking Breathing to Metabolism. Cell Rep. 2020, 33, 108358. [Google Scholar] [CrossRef]
- Neyens, D.; Zhao, H.; Huston, N.J.; Wayman, G.A.; Ritter, R.C.; Appleyard, S.M. Leptin Sensitizes NTS Neurons to Vagal Input by Increasing Postsynaptic NMDA Receptor Currents. J. Neurosci. Off. J. Soc. Neurosci. 2020, 40, 7054–7064. [Google Scholar] [CrossRef] [PubMed]
- Bassi, M.; Furuya, W.I.; Menani, J.V.; Colombari, D.S.A.; do Carmo, J.M.; da Silva, A.A.; Hall, J.E.; Moreira, T.S.; Wenker, I.C.; Mulkey, D.K.; et al. Leptin into the Ventrolateral Medulla Facilitates Chemorespiratory Response in Leptin-Deficient (Ob/Ob) Mice. Acta Physiol. Oxf. Engl. 2014, 211, 240–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Z.; Hao, Y.; Yu, H.; Shi, L.; Jing, X.; Zhang, X.; Liu, N.; Li, T.; Zhang, X.; Yuan, F. Disordered Leptin Signaling in the Retrotrapezoid Nucleus Is Associated with the Impaired Hypercapnic Ventilatory Response in Obesity. Life Sci. 2020, 257, 117994. [Google Scholar] [CrossRef] [PubMed]
- Zoccal, D.B.; Furuya, W.I.; Bassi, M.; Colombari, D.S.A.; Colombari, E. The Nucleus of the Solitary Tract and the Coordination of Respiratory and Sympathetic Activities. Front. Physiol. 2014, 5, 238. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, A.B.; Kirkness, J.P.; Smith, P.L.; Schneider, H.; Polotsky, M.; Richardson, R.A.; Hernandez, W.C.; Schwartz, A.R. Novel Whole Body Plethysmography System for the Continuous Characterization of Sleep and Breathing in a Mouse. J. Appl. Physiol. 2012, 112, 671–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Condos, R.; Norman, R.G.; Krishnasamy, I.; Peduzzi, N.; Goldring, R.M.; Rapoport, D.M. Flow Limitation as a Noninvasive Assessment of Residual Upper-Airway Resistance during Continuous Positive Airway Pressure Therapy of Obstructive Sleep Apnea. Am. J. Respir. Crit. Care Med. 1994, 150, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, A.R.; Brower, R.; Smith, P.; Wise, R.; Permutt, S. Mechanism of Inspiratory Flow Limitation in the Isolated Canine Upper Airway during Neuromuscular Blockade. Am. Rev. Respir. Dis. 1989, 139, A79. [Google Scholar]
- Gold, A.R.; Schwartz, A.R. The Pharyngeal Critical Pressure. The Whys and Hows of Using Nasal Continuous Positive Airway Pressure Diagnostically. Chest 1996, 110, 1077–1088. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, A.R.; Smith, P.L.; Wise, R.A.; Gold, A.R.; Permutt, S. Induction of Upper Airway Occlusion in Sleeping Individuals with Subatmospheric Nasal Pressure. J. Appl. Physiol. 1988, 64, 535–542. [Google Scholar] [CrossRef]
- Smith, P.L.; Wise, R.A.; Gold, A.R.; Schwartz, A.R.; Permutt, S. Upper Airway Pressure-Flow Relationships in Obstructive Sleep Apnea. J. Appl. Physiol. 1988, 64, 789–795. [Google Scholar] [CrossRef]
- Schwartz, A.R.; Smith, P.L.; Wise, R.A.; Bankman, I.; Permutt, S. Effect of Positive Nasal Pressure on Upper Airway Pressure-Flow Relationships. J. Appl. Physiol. 1989, 66, 1626–1634. [Google Scholar] [CrossRef] [PubMed]
- Bonaventura, J.; Eldridge, M.A.G.; Hu, F.; Gomez, J.L.; Sanchez-Soto, M.; Abramyan, A.M.; Lam, S.; Boehm, M.A.; Ruiz, C.; Farrell, M.R.; et al. High-Potency Ligands for DREADD Imaging and Activation in Rodents and Monkeys. Nat. Commun. 2019, 10, 4627. [Google Scholar] [CrossRef] [Green Version]
- Fleury Curado, T.; Pho, H.; Freire, C.; Amorim, M.R.; Bonaventura, J.; Kim, L.J.; Lee, R.; Cabassa, M.E.; Streeter, S.R.; Branco, L.G.; et al. Designer Receptors Exclusively Activated by Designer Drugs Approach to Treatment of Sleep-Disordered Breathing. Am. J. Respir. Crit. Care Med. 2021, 203, 102–110. [Google Scholar] [CrossRef]
- Dampney, R.A. Functional Organization of Central Pathways Regulating the Cardiovascular System. Physiol. Rev. 1994, 74, 323–364. [Google Scholar] [CrossRef] [PubMed]
- Machado, B.H. Neurotransmission of the Cardiovascular Reflexes in the Nucleus Tractus Solitarii of Awake Rats. Ann. N. Y. Acad. Sci. 2001, 940, 179–196. [Google Scholar] [CrossRef]
- Nichols, N.L.; Hartzler, L.K.; Conrad, S.C.; Dean, J.B.; Putnam, R.W. Intrinsic Chemosensitivity of Individual Nucleus Tractus Solitarius (NTS) and Locus Coeruleus (LC) Neurons from Neonatal Rats. Adv. Exp. Med. Biol. 2008, 605, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Mercer, J.G.; Moar, K.M.; Hoggard, N. Localization of Leptin Receptor (Ob-R) Messenger Ribonucleic Acid in the Rodent Hindbrain. Endocrinology 1998, 139, 29–34. [Google Scholar] [CrossRef]
- Hosoi, T.; Kawagishi, T.; Okuma, Y.; Tanaka, J.; Nomura, Y. Brain Stem Is a Direct Target for Leptin’s Action in the Central Nervous System. Endocrinology 2002, 143, 3498–3504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassi, M.; Giusti, H.; Leite, C.M.; Anselmo-Franci, J.A.; do Carmo, J.M.; da Silva, A.A.; Hall, J.E.; Colombari, E.; Glass, M.L. Central Leptin Replacement Enhances Chemorespiratory Responses in Leptin-Deficient Mice Independent of Changes in Body Weight. Pflugers Arch. 2012, 464, 145–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassi, M.; Furuya, W.I.; Zoccal, D.B.; Menani, J.V.; Colombari, D.S.A.; Mulkey, D.K.; Colombari, E. Facilitation of Breathing by Leptin Effects in the Central Nervous System. J. Physiol. 2016, 594, 1617–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prabhakar, N.R. Sensing Hypoxia: Physiology, Genetics and Epigenetics. J. Physiol. 2013, 591, 2245–2257. [Google Scholar] [CrossRef]
- Ortega-Sáenz, P.; López-Barneo, J. Physiology of the Carotid Body: From Molecules to Disease. Annu. Rev. Physiol. 2020, 82, 127–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caballero-Eraso, C.; Shin, M.-K.; Pho, H.; Kim, L.J.; Pichard, L.E.; Wu, Z.-J.; Gu, C.; Berger, S.; Pham, L.; Yeung, H.-Y.; et al. Leptin Acts in the Carotid Bodies to Increase Minute Ventilation during Wakefulness and Sleep and Augment the Hypoxic Ventilatory Response. J. Physiol. 2019, 597, 151–172. [Google Scholar] [CrossRef] [Green Version]
- Shin, M.-K.; Kim, L.J.; Caballero-Eraso, C.; Polotsky, V.Y. Experimental Approach to Examine Leptin Signaling in the Carotid Bodies and Its Effects on Control of Breathing. JoVE J. Vis. Exp. 2019, e60298. [Google Scholar] [CrossRef]
- Shin, M.-K.; Eraso, C.C.; Mu, Y.-P.; Gu, C.; Yeung, B.H.Y.; Kim, L.J.; Liu, X.-R.; Wu, Z.-J.; Paudel, O.; Pichard, L.E.; et al. Leptin Induces Hypertension Acting on Transient Receptor Potential Melastatin 7 Channel in the Carotid Body. Circ. Res. 2019, 125, 989–1002. [Google Scholar] [CrossRef] [PubMed]
- Ciriello, J.; Caverson, M.M. Carotid Chemoreceptor Afferent Projections to Leptin Receptor Containing Neurons in Nucleus of the Solitary Tract. Peptides 2014, 58, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Freire, C.; Pho, H.; Bevans-Fonti, S.; Sennes, L.U.; Polotsky, V.Y. Intranasal Leptin Improves Survival after Opioid Overdose in a Mouse Model. J. Transl. Med. 2021, 19, 134. [Google Scholar] [CrossRef]
- Williams, K.W.; Smith, B.N. Rapid Inhibition of Neural Excitability in the Nucleus Tractus Solitarii by Leptin: Implications for Ingestive Behaviour. J. Physiol. 2006, 573, 395–412. [Google Scholar] [CrossRef]
- Gastaut, H.; Tassinari, C.A.; Duron, B. Polygraphic Study of the Episodic Diurnal and Nocturnal (Hypnic and Respiratory) Manifestations of the Pickwick Syndrome. Brain Res. 1966, 1, 167–186. [Google Scholar] [CrossRef]
- Patil, S.P.; Schneider, H.; Marx, J.J.; Gladmon, E.; Schwartz, A.R.; Smith, P.L. Neuromechanical Control of Upper Airway Patency during Sleep. J. Appl. Physiol. 2007, 102, 547–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwab, R.J.; Wang, S.H.; Verbraecken, J.; Vanderveken, O.M.; Van de Heyning, P.; Vos, W.G.; DeBacker, J.W.; Keenan, B.T.; Ni, Q.; DeBacker, W. Anatomic Predictors of Response and Mechanism of Action of Upper Airway Stimulation Therapy in Patients with Obstructive Sleep Apnea. Sleep 2018, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleury Curado, T.; Oliven, A.; Sennes, L.U.; Polotsky, V.Y.; Eisele, D.; Schwartz, A.R. Neurostimulation Treatment of OSA. Chest 2018, 154, 1435–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polotsky, M.; Elsayed-Ahmed, A.S.; Pichard, L.E.; Richardson, R.A.; Smith, P.L.; Schneider, H.; Kirkness, J.P.; Polotsky, V.Y.; Schwartz, A.R. Effect of Age and Weight on Upper Airway Function in a Mouse Model. J. Appl. Physiol. 2011, 111, 696–703. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amorim, M.R.; Dergacheva, O.; Fleury-Curado, T.; Pho, H.; Freire, C.; Mendelowitz, D.; Branco, L.G.S.; Polotsky, V.Y. The Effect of DREADD Activation of Leptin Receptor Positive Neurons in the Nucleus of the Solitary Tract on Sleep Disordered Breathing. Int. J. Mol. Sci. 2021, 22, 6742. https://doi.org/10.3390/ijms22136742
Amorim MR, Dergacheva O, Fleury-Curado T, Pho H, Freire C, Mendelowitz D, Branco LGS, Polotsky VY. The Effect of DREADD Activation of Leptin Receptor Positive Neurons in the Nucleus of the Solitary Tract on Sleep Disordered Breathing. International Journal of Molecular Sciences. 2021; 22(13):6742. https://doi.org/10.3390/ijms22136742
Chicago/Turabian StyleAmorim, Mateus R., Olga Dergacheva, Thomaz Fleury-Curado, Huy Pho, Carla Freire, David Mendelowitz, Luiz G. S. Branco, and Vsevolod Y. Polotsky. 2021. "The Effect of DREADD Activation of Leptin Receptor Positive Neurons in the Nucleus of the Solitary Tract on Sleep Disordered Breathing" International Journal of Molecular Sciences 22, no. 13: 6742. https://doi.org/10.3390/ijms22136742