Lipid Rafts and Dopamine Receptor Signaling


1
Department of Medicine, Einstein Medical Center Philadelphia, Philadelphia, PA 19141, USA
2
Division of Renal Diseases & Hypertension, Department of Medicine, The George Washington University School of Medicine & Health Sciences, Washington, DC 20052, USA
3
Department of Pharmacology/Physiology, The George Washington University School of Medicine & Health Sciences, Washington, DC 20052, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(23), 8909; https://doi.org/10.3390/ijms21238909
Submission received: 30 October 2020
/
Revised: 18 November 2020
/
Accepted: 20 November 2020
/
Published: 24 November 2020
(This article belongs to the Special Issue Raft Microdomains and Cell Signaling)
Abstract
:The renal dopaminergic system has been identified as a modulator of sodium balance and blood pressure. According to the Centers for Disease Control and Prevention, in 2018 in the United States, almost half a million deaths included hypertension as a primary or contributing cause. Renal dopamine receptors, members of the G protein-coupled receptor family, are divided in two groups: D1-like receptors that act to keep the blood pressure in the normal range, and D2-like receptors with a variable effect on blood pressure, depending on volume status. The renal dopamine receptor function is regulated, in part, by its expression in microdomains in the plasma membrane. Lipid rafts form platforms within the plasma membrane for the organization and dynamic contact of molecules involved in numerous cellular processes such as ligand binding, membrane sorting, effector specificity, and signal transduction. Understanding all the components of lipid rafts, their interaction with renal dopamine receptors, and their signaling process offers an opportunity to unravel potential treatment targets that could halt the progression of hypertension, chronic kidney disease (CKD), and their complications.
1. Introduction
The classical definition of a cell membrane is a semipermeable membrane, composed of a lipid bilayer mixed with proteins, that surrounds the cytoplasm and nucleoplasm. The cell membrane demarcates the inner from the outer cellular environment. The phospholipids and sphingolipids in the cell membrane function as building blocks because of their aggregative properties. These phospholipids consist of a hydrophilic head and hydrophobic tails. The hydrophobic tails interact with each other whereas the hydrophilic heads face the inner cellular and extracellular space. In 1997, the concept of rigid lipids and protein mixture changed when the formalized hypothesis on lipid rafts was published as a concept paper on “functional rafts” [1]. Further research has helped in the understanding of the compartmentalization of cell membranes into lipid and non-lipid raft microdomains [2]. Many receptors are synthesized intracellularly and targeted to these microdomains, which assist receptors in numerous cellular processes including, but not limited to, ligand affinity and cellular signal transduction. Several studies involving dopamine receptors have shown the delicate interaction between dopamine receptors and lipid rafts, as well as the need of functional lipid rafts so that dopamine receptors can operate properly [2].
The renal dopaminergic system plays a key role in the control of blood pressure, in part by regulating renal sodium transport [3]. In 2018, almost half a million deaths included hypertension as a primary or contributing cause. Hypertension is caused, in part, by expansion of the extracellular fluid, because of sodium retention due to an inability to excrete a sodium load [3,4,5]. For this reason, the study of lipid rafts and dopamine receptors could offer therapeutic targets related to essential hypertension. The activation of dopamine receptors has been shown to participate in the reduction of reactive oxygen species production and inflammation, which cause the progression of chronic kidney disease (CKD) [4,5]. Thus, further research on this topic can open new targets to stop the progression of hypertension, CKD, and their complications.
2. Definition of Lipid Rafts, Structure, Function, and Associated Components
Lipid rafts (Figure 1) are dynamic and shifting assemblies of sphingolipids, cholesterol, glycosphingolipids, and proteins forming platforms or lipid microdomains for the organization and dynamic contact of molecules involved in several molecular and cellular processes such as ligand affinity, effector specificity, signal transduction, membrane sorting, and receptor trafficking and recycling. Cholesterol molecules rest between the hydrophobic tails of phospholipids; next to them, many proteins serve as anchor units for other proteins. The organization of certain groups of proteins allows optimal function, as well as specification of cellular signal transmission by granting effective interaction between proteins and avoiding interference of opposite signaling pathways [2,6]. Lipid rafts are located in cell surface membranes, as well as in intracellular membranes. These allow the coordination of cellular processes, which in turn affect bioactivity of many receptors and the cell itself [2].
Many proteins are associated with lipid rafts; one of the best characterized proteins are the caveolins. These form caveolae (“little caves”). Caveolae (Figure 2) are scaffolding proteins that assemble within lipid raft domains to form flask-shaped invaginations of the plasma membrane. These platforms allow protein–protein, lipid–lipid, and protein–lipid interactions and regulate numerous cellular processes (signal transduction, receptor trafficking, recycling, etc.). There are three caveolin isoforms (Cav1, Cav2, and Cav3). However, Cav1 is the isoform that is present in most tissues and is enough for the formation of caveolae [7].
The function of surface membrane receptors is determined by their intracellular trafficking pathway. These receptors are produced intracellularly and their trafficking to the plasma membrane is needed for the attachment to the extracellular agonist or antagonist. Agonist-induced activation of the receptor is followed by its subsequent internalization and re-insertion to the plasma membrane (desensitization and resensitization of the receptor), ultimately controlling the function and activity of the receptor [7,8,9,10,11].
G protein-coupled receptors (GPCRs) are the largest superfamily of mammalian surface membrane receptors that work as signaling proteins. These are involved in diverse physiological processes by binding to ligands and transducing intracellular signaling pathways via activation of G proteins [9,10,11], i.e., G protein subunits α, β, and γ. The structure of GPCRs features the presence of seven transmembrane α-helical molecules, which are connected by three extracellular and three intracellular loops [10].
Surface membrane protein receptors, including GPCRs, undergo a trafficking process with constant co-translational and post-translational changes before arriving at the plasma membrane. As with many cellular proteins, GPCRs, following their synthesis, reside in the endoplasmic reticulum (ER) where they undergo assembly, folding, and packaging. The GPCRs temporarily reside in endoplasmic reticulum-derived COPII (coat protein complex II) transport vesicles, before exiting the ER into the Golgi apparatus. There, the GPCRs undergo further modifications, such as the S-palmitoylation of cysteine residues. At the Golgi edge, the GPCRs are boxed again into vesicles, into the endosomal pathway, which targets the receptors to the plasma membrane. In the plasma membrane, the GPCRs mature and become functional. Some subsets of GPCRs are targeted to specific regions of the plasma membrane, such as the lipid rafts [11].
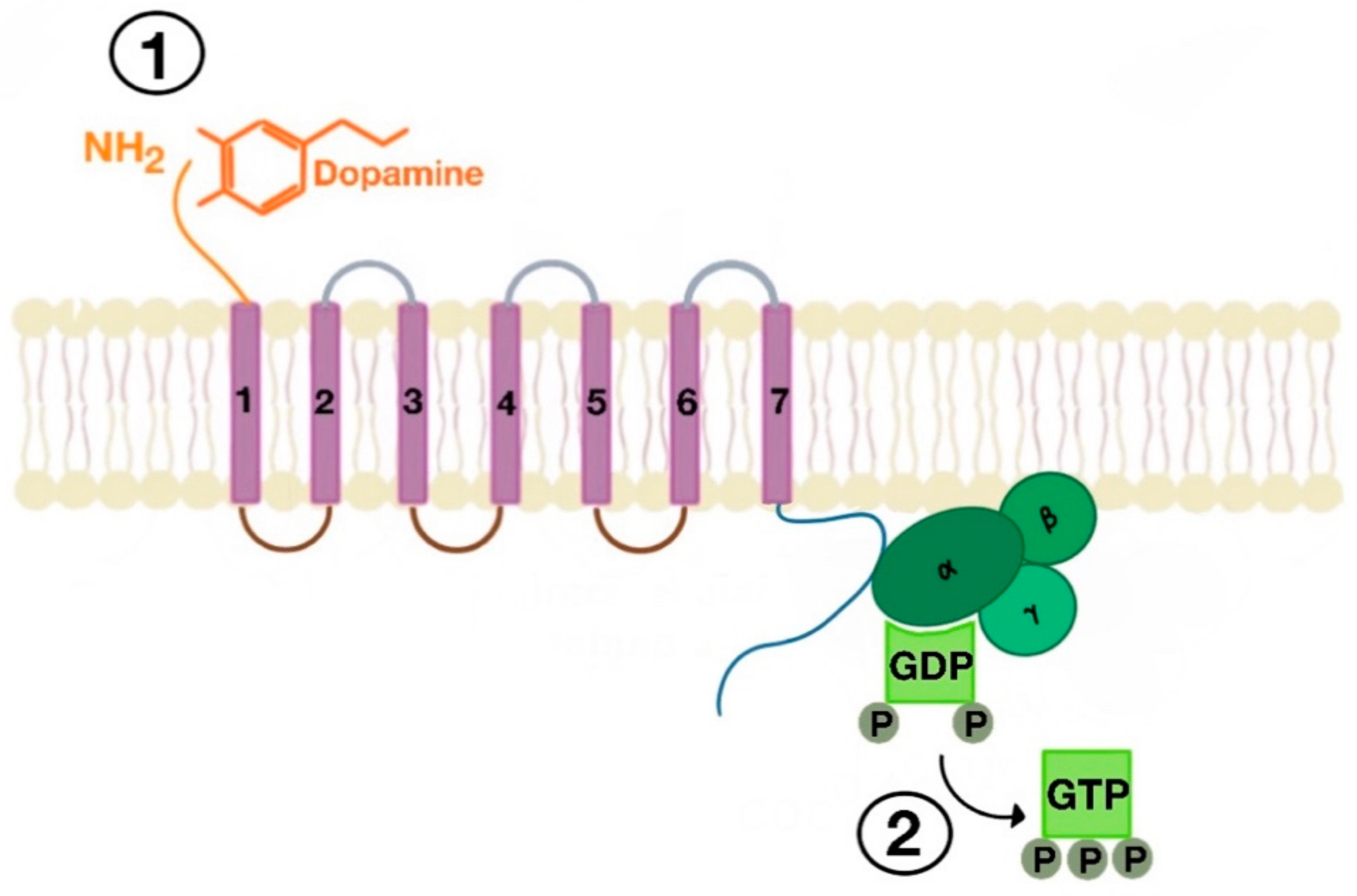
There are many motifs of proteins in the GPCRs that interact with the plasma membrane and its associated cholesterol. These motifs carry a cholesterol recognition/interaction amino acid consensus or CRAC motif. The CRAC motif helps anchor the Cav isoforms to the plasma membrane [12] as well as other plasma proteins, including GPCRs. Once the GPCR is targeted to a lipid raft, it can be successfully stimulated by interacting with its ligand (Figure 3). The agonist occupation of GPCRs, as in dopamine receptors, results in a conformational change promoting the exchange of GDP (guanosine diphosphate) to GTP (guanosine triphosphate) on the Gα subunit of the G protein, leading to two actions (Figure 4). First, the uncoupling of the G proteins from the GPCR, and second, the uncoupling of the Gα subunit from the Gβγ subunit. Finally, the Gα subunit stimulates or inactivates downstream signaling processes, depending on its conformational and functional characterization, Gαs being stimulatory or Gαi being inhibitory of adenylyl cyclase activity. The remaining Gβγ subunit activates GPCR kinases (GRKs), which phosphorylate amino acids in the third intracellular loop of the transmembrane segments and C-terminal tail of the receptor, leading to the activation of cytosolic proteins known as arrestins [13,14] (Figure 5). The activation of GRKs and β-arrestins ensures the removal of the GPCR from the cell surface, leading to its desensitization.
The β-arrestins, originally discovered for their inhibitory part in some receptor signaling pathways, have been proven to work as critical controllers of agonist-induced endocytosis and ubiquitination of plasma receptors, including GPCRs. Therefore, β-arrestins play a pivotal role in the trafficking path of endocytosed GPCRs [13,14].
The β-arrestins direct sequestration of GPCRs via a dynamin-dependent, clathrin-mediated endocytosis. This process takes effect through adaptor protein 2 (AP2) complexes, which in turn begin clathrin-coated pit assembly (Figure 6). As soon as the GPCR is internalized, it is packaged in early endosome (Figure 7), where the GPCR is dephosphorylated by protein phosphatase 2A, in the case of the D1R [14]. Then, sorting nexins (SNXs) distribute the GPCR to different destinations [15]. From the endosome, the receptor is either sent to fast recycling endosome and subsequently to the lipid raft in the plasma membrane, or to lysosomes (Figure 8) for eventual degradation.
The SNXs are a diverse group of membrane-linked phosphoinositide-binding proteins that regulate cellular protein trafficking by controlling all aspects of the endocytic pathway, including endocytosis, endosomal sorting, and endosomal signaling [16]. The SNX either sorts the endocytosed and dephosphorylated/resensitized GPCR into the fast recycling endosomes, targeting it to lipid rafts (Figure 9), or send such receptor to late endosomes, targeting it to lysosomes, inducing the inevitable degradation of the GPCR [17].
3. Definition of Dopamine and Classification of Dopamine Receptors
Lipid rafts are involved in the regulation of dopamine receptors. Dopamine is a catecholamine that functions as a precursor of norepinephrine and epinephrine. However, during the past decades, it has been identified as a modulator of sodium balance and blood pressure by means of a renal dopaminergic system. Dopamine interacts with GPCRs that are classified into five genetically distinct types [3,4,5,18,19,20,21]: D1R, D2R, D3R, D4R, and D5R.
The dopamine receptors listed above are divided in two groups, D1-like and D2-like receptors, all of which are expressed in the kidney (Table 1). The D1-like receptors are coupled to G proteins Gs and Gq to stimulate adenylyl cyclase and phospholipase C activity, respectively; D1R and D5R belong to this group. The D2-like receptors are coupled to Gi and Go to inhibit adenylyl cyclase activity; D2R, D3R, and D4R belong to this group [3,4,5,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38].
Dopamine receptors, like other GPCRs, have seven transmembrane domains, linked by three intracellular loops, three extracellular loops, an extracellular amino terminus, and an intracellular cytoplasmic C terminal tail [20,21]. Members of the same family of dopamine receptors (D1-like and D2-like) display considerable structural homology in terms of their transmembrane domains, NH2—terminal and COOH terminal domains [20,21].
However, the D1-like receptors have a short third intracellular loop whereas the D2-like receptors have a long third intracellular loop [20,21]. The NH2-terminal domain has the same number of amino acids in all the D1-like and D2-like receptors and contains a variable number of consensus N-glycosylation sites (Table 2) [8,20,21].
In the D1-like receptors, the COOH terminus is about seven times longer than that in D2-like receptors (Figure 10). The COOH terminus has abundant serine and threonine residues and contains a cysteine residue that is present in all GPCRs. In the D1-like receptors, the location of the cysteine residue is near the start of the COOH terminus. However, for the D2-like receptors, the COOH terminus ends with a cysteine residue [20].
4. Function of Renal D1-Like and D2-Like Receptors
A dysfunction in the production of intrarenal dopamine or dopamine receptor signaling can predispose to hypertension, with or without salt sensitivity that is related, in part, to dysregulation of the renin–angiotensin system [3,4,5,18,19]. Additionally, several studies indicate that renal dopamine and its receptors play a role in the reduction of reactive oxygen species production, inflammation, and the progression of CKD [3,5,19]. As the receptor signaling system depends on lipid rafts, failure to assemble and maintain the lipid raft, as well as to target the dopamine receptors to lipid rafts, would result in the aforementioned pathological states.
Dopamine receptors are important in establishing normal sodium balance and normal blood pressure under conditions of euvolemia and moderate volume expansion. Renal dopamine D1-like receptors cause renal vasodilation and natriuresis, the latter being due to inhibition of ion transport at the apical and basolateral membranes of renal tubules [3,4,5,18,19,22,23]. The natriuretic effect of dopamine is primarily exerted through D1R and D5R by inhibiting both the influx and efflux of Na+ in renal epithelial cells. This is accomplished by inhibiting multiple Na+ cotransporters, exchangers, and pump along the nephron, resulting in an increase in Na+ excretion [3,4,5,18,19,22,23].
Renal D2-like receptors also participate in the inhibition of ion transport during conditions of euvolemia and moderate volume expansion [3,5,18,19,22,23]. D2R, D3R, and D4R (D2-like receptors) are also involved in the CNS regulation of blood pressure; post-synaptic D2-like receptors increase blood pressure, while presynaptic D2-like receptors (the predominant action) produce the opposite effect [3,23].
5. Signaling Pathway of Renal Dopamine Receptors
Similar to other GPCRs, the dopamine signaling pathway is divided in four stages: (1) activation of dopamine receptor, (2) interaction of receptor with heterotrimeric G proteins, (3) signal transduction through effector molecules, and (4) cellular response. As with other GPCRs, the dopamine receptors need to be internalized, endocytosed, and retargeted to the lipid raft to be functional again. These processes cause desensitization and resensitization.
As previously mentioned, D1-like and D2-like receptors belong to the family of membrane receptors called GPCRs. GPCRs exert their actions via heterotrimeric G protein subunits α, β, and γ.
5.1. D1-Like Receptor Signaling Pathway
D1-like receptors are coupled with Gαs and Gαq [3,4,5,18,19,20,21,22,23,24,25]. The D1-like receptors, coupled with Gs, activate adenylyl cyclase, which breaks down ATP into cAMP (cyclic adenosine monophosphate). cAMP leads to the activation of protein kinase A, which functions as an effector molecule that inhibits the Na+/H+ exchanger type 3 (NHE3) at the brush border side of the proximal tubule and thick ascending limb of Henle [3,5,18,19,20,21,22,23,24,25,26] (Figure 11). However, NHE3 can be directly inhibited by Gsα, independent of protein kinase A, at least in renal proximal tubule brush border membranes. In these membranes, β/γ dimers can stimulate NHE3 activity [26]. The D1R, but not D5R, can couple to Go [27]. By contrast, D5R but not D1R can couple to Gz and Gα 12/13 [28,29].
The D1-like receptors’ linkage to Gq activates phospholipase C, which in turn cleaves phosphatidylinositol 4,5-bisphosphate (lipid molecule found in the intracellular side of the cell plasma membrane, as well as in membranes of several intracellular organelles (C)) into DAG (diacylglycerol) and IP3 (inositol-1,4,5-trisphosphate). DAG activates protein kinase C (effector molecule) that ultimately inhibits Na+/K+-ATPase on the basolateral side of the nephron (Figure 12). The linkage of G protein subunits to D1-like receptors may be tissue-specific [30]. The effect of dopamine receptors on sodium transport is also tissue-specific. The D5R increases NHE3 activity in human embryonic kidney (HEK)293T cells [31].
5.2. D2-Like Receptor Signaling Pathway
The D2-like receptors, which consist of D2R, D3R, and D4R, couple to Gαi, which inhibits adenylyl cyclase and reduces cAMP production. D2-like receptors are also linked to Go [3,4,5,18,19,20,21,22,23,24]. D2-like receptors also inhibit calcium channel activity and modulate potassium channel activity [3,32,33,34]. D2R can couple to the same extent to Gαi and Gz but not to Gq11 or Gα12/13. There are two isoforms of D2R, the D2short and D2long; the latter is expressed in the kidney [35]. The D2short, via Rho A, can couple to phospholipase D [3,36]. In contrast to the other D2-like receptors, D3R linkage to Gαi is not robust and can actually be linked to Gαs and Gq11 [3,24,37,38].
6. Role of the D1-Like and D2-Like Receptors in Natriuresis and Antinatriuresis
Dopamine in the kidney plays a pivotal role in the inhibition of sodium, particularly in conditions of moderate sodium excess [3,4,5,18,19,22,24]. An increase in extracellular fluid increases renal tubular dopamine production and the converse decreases renal tubular dopamine production [3,39]. When extracellular fluid volume is decreased, the intravenous infusion of dopamine decreases sodium excretion, whereas when extracellular fluid volume is expanded, the intravenous infusion of dopamine increases sodium excretion [40]. Renal proximal tubule-selective deletion of dopamine synthesis impairs sodium excretion and increases blood pressure [19,41]. Long-term pharmacological blockade of dopamine receptor subtypes also decreases renal sodium excretion and increases blood pressure [3,18,19,22,24,42,43,44,45,46]. Moreover, inactivation of any dopamine receptor subtype gene in mice results in hypertension that may be salt-sensitive, depending on the genetic background [3,25,46]. The stimulation of D1-like receptors mediate natriuresis by inhibiting NHE3 (SLC9A3), sodium phosphate cotransporter (NaPi-IIa/SLC34A1 and NaPIIc/SLC34A3), Cl−/HCO3− exchanger (SLC26A6) at the apical membrane, and electrogenic Na/HCO3 cotransporter (NBCe2, SLC4A5 [47,48]) and Na+/K+-ATPase at the basolateral membrane [3,18,19,23,25,49,50,51,52,53,54,55]. Currently, there are no commercially available agonists that can differentiate between the subtypes of D1-like receptors D1R and D5R [3]. Stepholidine is a D1R agonist and D2R/D3R antagonist [56]. LE-PM436 is a selective D5R antagonist [57]. The D1R and D5R interact to inhibit NHE3 and Na+/K+/ATPase activity via both the adenylyl cyclase and phospholipase C pathways [58]. In conscious rats, the renal interstitial infusion of D1R-specific antisense oligodeoxynucleotides impairs sodium excretion, providing evidence of the importance of the D1R in the inhibition of renal tubular sodium transport [49].
The effect of D2-like receptors on renal sodium transport is variable, including no effect, attenuation of sodium excretion, and increase in sodium excretion that may be related to the state of extracellular fluid volume [3,40]. Bromocriptine, a D2-like receptor agonist (D2R = D3R > D4R), as well as serotonin agonist increased Na+/K+/ATPase activity in rat renal proximal tubules [59]. The D2R agonist LY171555 increased Na+/K+/ATPase activity in Ltk-11 cells heterologously expressing the D2long receptor [60]. By contrast, the D3R agonist PD128907 inhibited Na+/K+/ATPase activity in rat renal proximal tubules [61]. The D4R agonist PD168077 also decreased the ability of insulin to stimulate Na+/K+/ATPase activity in rat renal proximal tubule cells [62]. Sulpiride (D2R = D3R > D4R), a D2-like receptor antagonist, impaired dopamine-mediated natriuresis in euvolemic males [63]. The D3R agonists PD128907 and 7-OH-DPAT increased sodium excretion in rats chronically fed a high-salt diet [61,64]. It is possible that activation of D2-like receptors may cause antinatriuresis or natriuresis in volume-depleted and volume-expanded states, respectively [25,40]. The role of specific D2-like receptors in this process needs to be clarified.
7. Renal D1-Like Receptors and Lipid Rafts
Lipid rafts are important in organizing signal transduction cascades [1,2,6,7,8,9,12,15,16,65,66]. D1R partitions to lipid as well as non-lipid rafts in human renal proximal tubule cells expressing wild-type GRK4 [67,68,69,70] and interacts with the lipid raft protein Cav1 [67,68,71,72,73,74,75] in renal proximal tubule cells. Additionally, it interacts Cav2 in HEK293 cells expressing D1R [76]. By contrast, in the presence of increased GRK4 activity, which is characteristic of GRK4 variants [46,77,78,79,80,81,82,83,84,85,86], in human renal proximal tubule cells, or increased renal GRK4 expression in rodents, the D1R may partition mainly to non-lipid rafts [87]. The D5R, as with the D1R, also partitions to lipid as well as non-lipid rafts in human renal proximal tubule cells [68], but in rat renal proximal tubule cells, the D1R and D5R are mainly expressed in non-lipid rafts [87]. However, GRK2, not GRK4, regulates the D5R [31]. Disruption of lipid rafts in human renal proximal tubule cells with methyl-β-cyclodextrin (β-MCD) [88] blocks the fenoldopam (D1-like receptor (D1R and D5R) agonist)-stimulated cAMP production [67]. β-MCD removes caveolar domains very efficiently but also non-specifically depletes plasma membrane cholesterol. Kidney-restricted disruption of lipid rafts in rats impairs D1-like receptor-mediated natriuresis and increases blood pressure [72].
As aforementioned, the SNX family plays a pivotal role in the sorting and transport of GPCRs through a series of endosomal compartments [89,90,91]. The first 20 min of D1R desensitization is in part due to SNX5 [69], and also to SNX19 (unpublished), and thereafter GRK4, and, to a lesser extent, GRK2 [84]. SNX5 is important in the recycling of glycosylated D1R to the plasma membrane that requires the di-leucine motif (L344–L345) at the D1R C-terminus [8]. The mutation of two specific palmitoylation sites (C347 and Y218) of a CRAC motif resulted in the absence of D1R in lipid rafts [70]. Some of the recycled D1R has to be targeted to lipid rafts to be functional; this is not afforded by SNX5, probably because it is mainly expressed in non-lipid rafts. The recycling of D1R to lipid rafts is probably due to SNX19, which is normally exclusively found in the lipid raft [70]. For D1R to function, a critical amount of D1R has to be inserted in lipid rafts where adenylyl cyclase 6 is expressed; adenylyl cyclase 6 is activated to a greater extent by D1R than D5R [67]. Adenylyl cyclase isoforms 2, 3, 6, 7, and 9 are expressed in the renal proximal tubule [92]. Acute siRNA (small interfering RNA)-mediated, renal-restricted depletion of SNX5 decreases sodium excretion and further elevates the already increased blood pressure of spontaneously hypertensive rats [69]. SNX5 and D1R are also important in the normal expression of insulin receptor in human renal proximal tubule cells [93].
As indicated above, SNX19, which is found exclusively in lipid rafts, participates in the early desensitization of D1R in human renal proximal tubules. siRNA-mediated silencing of SNX19 in human renal proximal tubule cells impairs the ability of the D1R/D5R agonist fenoldopam to inhibit renal sodium transport. Renal-restricted siRNA-mediated depletion of SNX19 in mice decreases D1R expression and increases oxidative stress and blood pressure [70,94]. Lipid raft bestows protection against oxidative stress by maintaining NOX (NADPH oxidase) in the inactive state in human renal proximal tubule cells [68], the opposite of what is observed in normotensive rat renal proximal tubule cells [87], thus the need for caution in extrapolating non-human studies to human physiology, pathophysiology, or pharmacology.
SNX1 is required for D5R trafficking following agonist stimulation. It strongly binds to D5R, but not to D1R [95]. Absence of SNX1 blunts the cAMP production via D5R and suppression of renal sodium transport. Renal-restricted, siRNA-mediated depletion of SNX1 in mice decreases sodium excretion and increases blood pressure [96]. SNX1−/− mice also have increased blood pressure; the natriuretic effect of the D1R/D5R agonist fenoldopam is also impaired in these mice [96]. The impaired natriuretic effect of fenoldopam is associated with increased renal sodium transport by Na/K ATPase, as well as with other sodium cotransporters and exchangers. Interestingly, SNX1−/− mice have increased oxidative stress; oxidative stress being the cause or effect of hypertension [16,18,19,96,97]. D1R and D5R dysfunction or deficiency increases blood pressure, in part by an increase in oxidative stress [3,4,5,16,18,19,70,97,98,99].
8. Renal D2-Like Receptors and Lipid Rafts
Renal D2-like receptors are also found in lipid rafts. However, there are not many studies that have explored the role and interaction between lipid rafts microdomains and each renal D2-like receptor. In mouse striatum and in particular HEK293T cells, heterologously expressing the D2Rlong, the D2R is in plasma membrane microcompartments that do not correspond to detergent-resistant lipid rafts; β-MCD does not increase the detergent solubility of D2R [100]. In Chinese hamster ovary cells heterologously expressing the adenosine A2A and D2Rlong, following agonist stimulation, they co-internalize with Cav1, which is expressed in lipid rafts [101]. However, in the rat frontal cortex, endogenous D2Rs are distributed in cytoplasmic, detergent-soluble, and detergent-resistant fractions but not in buoyant fractions, as is the case for D1R [102]. GPCRs, such as D1-like and D2-like receptors, need to link to β-arrestin for their sequestration/internalization. However, dopamine D2R and D3R have different interactions with arrestins; the D3R exhibits less translocation via arrestins than the D2R. Additionally, arrestin 3 preferentially binds to D2R over D3R [103]. Haloperidol, an antagonist to D2R, D3R, and D4R, as well serotonin receptors, can inhibit cholesterol biosynthesis and redistribute flotillin-1 in plasma membranes of SH-SY5Y neuroblastoma cells [104].
The C-terminal tail of D2-like receptors is shorter than those in D1-like receptors [20]. The C-terminal tails of the D2-like receptors are similar in their structure. All of them contain cysteine residues that have been shown to undergo palmitoylation to anchor the cytoplasmic tail of these receptors [105]. The palmitoylation is relevant for the functioning of the D3R and D4R. Different studies performed on D1-like receptors showed that S-palmitoylation or palmitoylation of cysteine residues in the C-terminal tail of these GPCRs aids with their targeting to the lipid raft and subsequently with their proper functioning [8,70].
As with D1-like receptors, SNXs are also involved in the regulation of D2-like receptors. SNX25 is expressed in many tissues, including the kidney [106]. The overexpression of SNX25 amplifies the expression of D1R and D2R. Additionally, it augments signaling mediated by D2R and disrupts the endocytosis and recycling of D2R [106]. However, it does not affect D1R desensitization [106,107]. SNX27 is involved in D2R activation of G protein-gated inwardly rectifying channels in the brain [108].
9. Conclusions
In summary, lipid rafts and microcompartments in lipid rafts are important in the function of GPCRs, including dopamine receptors. The dopamine receptor family is divided in two groups, all of which are expressed in the kidney, D1-like receptors (D1R and D5R) and D2-like receptors (D2R, D3R, and D4R). As the dopamine receptor signaling system depends on lipid rafts, failure to assemble and maintain the lipid raft, as well as to target the dopamine receptors to lipid rafts, results in impaired natriuresis and increased production of reactive oxygen species and inflammation, leading to essential hypertension and progression of CKD. D1-like receptors are coupled with Gαs and Gαq and D2-like receptors couple to Gαi and to Go [3,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38]. The SNX family regulates the endocytosis, endosomal sorting, and endosomal signaling of dopamine receptors, targeting them to plasma membrane microdomains [89,90,91]. CRAC motif helps anchor the Cav isoforms and dopamine receptors to the plasma membrane [12,70,109]. S-palmitoylation or palmitoylation of cysteine residues in the C-terminal tail of dopamine receptors aids with their targeting to lipid rafts [8,70]. Agonist-induced activation of the receptor is followed by its subsequent internalization and re-insertion to the plasma membrane (desensitization and resensitization of the receptor), therefore restarting the dopamine receptor cycle. Further research on this topic can open new targets to stop the progression of hypertension and CKD, and their complications.
Author Contributions
All authors have read and agreed to the published version of the manuscript. V.J.M. and A.C.T. conceived the idea based on 1R01DK119652; V.J.M. wrote the manuscript which was edited by A.C.T., L.D.A. and P.A.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Institutes of Health, USA, 5R01DK039308, 1R01DK119652, and 5P01HL074940.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Simons, K.; Ikonen, E. How cells handle cholesterol. Science 2000, 290, 1721–1726. [Google Scholar] [CrossRef] [Green Version]
- Villar, V.A.; Cuevas, S.; Zheng, X.; Jose, P.A. Localization and signaling of GPCRs in lipid rafts. Methods Cell Biol. 2016, 132, 3–23. [Google Scholar] [CrossRef]
- Armando, I.; Villar, V.A.; Jose, P.A. Dopamine and renal function and blood pressure regulation. Compr. Physiol. 2011, 1, 1075–1117. [Google Scholar] [CrossRef]
- Zeng, C.; Felder, R.A.; Jose, P.A. A new approach for treatment of hypertension: modifying D1 dopamine receptor function. Cardiovasc. Hematol. Agents Med. Chem. 2006, 4, 369–377. [Google Scholar] [CrossRef]
- Wang, X.; Villar, V.A.; Armando, I.; Eisner, G.M.; Felder, R.A.; Jose, P.A. Dopamine, kidney, and hypertension: Studies in dopamine receptor knockout mice. Pediatr. Nephrol. 2008, 23, 2131–2146. [Google Scholar] [CrossRef]
- Lingwood, D.; Simons, K. Lipid rafts as a membrane-organizing principle. Science 2010, 327, 46–50. [Google Scholar] [CrossRef] [Green Version]
- Busija, A.R.; Patel, H.H.; Insel, P.A. Caveolins and cavins in the trafficking, maturation, and degradation of caveolae: Implications for cell physiology. Am. J. Physiol. Cell Physiol. 2017, 312, C459–C477. [Google Scholar] [CrossRef]
- Guo, Y.; Jose, P.A. C-terminal di-leucine motif of dopamine D1 receptor plays an important role in its plasma membrane trafficking. PLoS ONE 2011, 6, e29204. [Google Scholar] [CrossRef] [Green Version]
- Wootten, D.; Christopoulos, A.; Marti-Solano, M.; Babu, M.M.; Sexton, P.M. Mechanisms of signalling and biased agonism in G protein-coupled receptors. Nat. Rev. Mol. Cell Biol 2018, 19, 638–653. [Google Scholar] [CrossRef]
- Rosenbaum, D.M.; Rasmussen, S.G.; Kobilka, B.K. The structure and function of G-protein-coupled receptors. Nature 2009, 459, 356–363. [Google Scholar] [CrossRef] [Green Version]
- Drake, M.T.; Shenoy, S.K.; Lefkowitz, R.J. Trafficking of G protein-coupled receptors. Circ. Res. 2006, 99, 570–582. [Google Scholar] [CrossRef]
- Jafurulla, M.; Tiwari, S.; Chattopadhyay, A. Identification of cholesterol recognition amino acid consensus (CRAC) motif in G-protein coupled receptors. Biochem. Biophys. Res. Commun. 2011, 404, 569–573. [Google Scholar] [CrossRef]
- Jean-Charles, P.Y.; Kaur, S.; Shenoy, S.K. G Protein-Coupled Receptor Signaling Through β-Arrestin-Dependent Mechanisms. J. Cardiovasc. Pharmacol. 2017, 70, 142–158. [Google Scholar] [CrossRef]
- Lefkowitz, R.J. G protein-coupled receptors. III. New roles for receptor kinases and beta-arrestins in receptor signaling and desensitization. J. Biol. Chem. 1998, 273, 18677–18680. [Google Scholar] [CrossRef] [Green Version]
- Worby, C.A.; Dixon, J.E. Sorting out the cellular functions of sorting nexins. Nat. Rev. Mol. Cell Biol. 2002, 3, 919–931. [Google Scholar] [CrossRef]
- Yang, J.; Villar, V.A.M.; Rozyyev, S.; Jose, P.A.; Zeng, C. The emerging role of sorting nexins in cardiovascular diseases. Clin. Sci. 2019, 133, 723–737. [Google Scholar] [CrossRef] [Green Version]
- Von Zastrow, M. Mechanisms regulating membrane trafficking of G protein-coupled receptors in the endocytic pathway. Life Sci. 2003, 74, 217–224. [Google Scholar] [CrossRef]
- Banday, A.A.; Lokhandwala, M.F. Dopamine receptors and hypertension. Curr. Hypertens. Rep. 2008, 10, 268–275. [Google Scholar] [CrossRef]
- Zhang, M.Z.; Harris, R.C. Antihypertensive mechanisms of intra-renal dopamine. Curr. Opin. Nephrol. Hypertens. 2015, 24, 117–122. [Google Scholar] [CrossRef] [Green Version]
- Missale, C.; Nash, S.R.; Robinson, S.W.; Jaber, M.; Caron, M.G. Dopamine receptors: From structure to function. Physiol. Rev. 1998, 78, 189–225. [Google Scholar] [CrossRef] [Green Version]
- Beaulieu, J.M.; Gainetdinov, R.R. The physiology, signaling, and pharmacology of dopamine receptors. Pharmacol. Rev. 2011, 63, 182–217. [Google Scholar] [CrossRef] [Green Version]
- Jose, P.A.; Eisner, G.M.; Felder, R.A. Role of dopamine receptors in the kidney in the regulation of blood pressure. Curr. Opin. Nephrol. Hypertens. 2002, 11, 87–92. [Google Scholar] [CrossRef]
- Jose, P.A.; Eisner, G.M.; Felder, R.A. Role of dopamine in the pathogenesis of hypertension. Clin. Exp. Pharmacol. Physiol. Suppl. 1999, 26, S10–S13. [Google Scholar]
- Preto, A.J.; Barreto, C.A.V.; Baptista, S.J.; Almeida, J.G.; Lemos, A.; Melo, A.; Cordeiro, M.N.D.S.; Kurkcuoglu, Z.; Melo, R.; Moreira, I.S. Understanding the Binding Specificity of G-Protein Coupled Receptors toward G-Proteins and Arrestins: Application to the Dopamine Receptor Family. J. Chem. Inf. Model. 2020, 60, 3969–3984. [Google Scholar] [CrossRef]
- Hussain, T.; Lokhandwala, M.F. Renal dopamine receptors and hypertension. Exp. Biol. Med. 2003, 228, 134–142. [Google Scholar] [CrossRef]
- Albrecht, F.E.; Xu, J.; Moe, O.W.; Hopfer, U.; Simonds, W.F.; Orlowski, J.; Jose, P.A. Regulation of NHE3 activity by G protein subunits in renal brush-border membranes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 278, R1064–R1073. [Google Scholar] [CrossRef]
- Kimura, K.; White, B.H.; Sidhu, A. Coupling of human D-1 dopamine receptors to different guanine nucleotide binding proteins. Evidence that D-1 dopamine receptors can couple to both Gs and G(o). J. Biol. Chem. 1995, 270, 14672–14678. [Google Scholar] [CrossRef] [Green Version]
- Sidhu, A.; Kimura, K.; Uh, M.; White, B.H.; Patel, S. Multiple coupling of human D5 dopamine receptors to guanine nucleotide binding proteins Gs and Gz. J. Neurochem. 1998, 70, 2459–2467. [Google Scholar] [CrossRef]
- Zheng, S.; Yu, P.; Zeng, C.; Wang, Z.; Yang, Z.; Andrews, P.M.; Felder, R.A.; Jose, P.A. Galpha12- and Galpha13-protein subunit linkage of D5 dopamine receptors in the nephron. Hypertension 2003, 41, 604–610. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.Q.; Wang, H.Y.; Friedman, E. Stimulated D(1) dopamine receptors couple to multiple Galpha proteins in different brain regions. J. Neurochem. 2001, 78, 981–990. [Google Scholar] [CrossRef]
- Roosterman, D. Agonist-dependent and -independent dopamine-1-like receptor signalling differentially regulates downstream effectors. FEBS J. 2014, 281, 4792–4804. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.X.; Monsma, F.J.; Sibley, D.R.; Chiodo, L.A. D2L, D2S, and D3 dopamine receptors stably transfected into NG108-15 cells couple to a voltage-dependent potassium current via distinct G protein mechanisms. Synapse 1996, 24, 156–164. [Google Scholar] [CrossRef]
- Werner, P.; Hussy, N.; Buell, G.; Jones, K.A.; North, R.A. D2, D3, and D4 dopamine receptors couple to G protein-regulated potassium channels in Xenopus oocytes. Mol. Pharmacol. 1996, 49, 656–661. [Google Scholar] [PubMed]
- Momiyama, T.; Koga, E. Dopamine D(2)-like receptors selectively block N-type Ca(2+) channels to reduce GABA release onto rat striatal cholinergic interneurones. J. Physiol. 2001, 533, 479–492. [Google Scholar] [CrossRef]
- Gao, D.Q.; Canessa, L.M.; Mouradian, M.M.; Jose, P.A. Expression of the D2 subfamily of dopamine receptor genes in kidney. Am. J. Physiol. 1994, 266, F646–F650. [Google Scholar] [CrossRef]
- Senogles, S.E. D2s dopamine receptor mediates phospholipase D and antiproliferation. Mol. Cell. Endocrinol. 2003, 209, 61–69. [Google Scholar] [CrossRef]
- Obadiah, J.; Avidor-Reiss, T.; Fishburn, C.S.; Carmon, S.; Bayewitch, M.; Vogel, Z.; Fuchs, S.; Levavi-Sivan, B. Adenylyl cyclase interaction with the D2 dopamine receptor family; differential coupling to Gi, Gz, and Gs. Cell. Mol. Neurobiol. 1999, 19, 653–664. [Google Scholar] [CrossRef]
- Newman-Tancredi, A.; Cussac, D.; Audinot, V.; Pasteau, V.; Gavaudan, S.; Millan, M.J. G protein activation by human dopamine D3 receptors in high-expressing Chinese hamster ovary cells: A guanosine-5’-O-(3-[35S]thio)- triphosphate binding and antibody study. Mol. Pharmacol. 1999, 55, 564–574. [Google Scholar]
- Du, Z.; Yan, Q.; Wan, L.; Weinbaum, S.; Weinstein, A.M.; Wang, T. Regulation of glomerulotubular balance. I. Impact of dopamine on flow-dependent transport. Am. J. Physiol. Ren. Physiol. 2012, 303, F386–F395. [Google Scholar] [CrossRef] [Green Version]
- Agnoli, G.C.; Cacciari, M.; Garutti, C.; Ikonomu, E.; Lenzi, P.; Marchetti, G. Effects of extracellular fluid volume changes on renal response to low-dose dopamine infusion in normal women. Clin. Physiol. 1987, 7, 465–479. [Google Scholar] [CrossRef]
- Zhang, M.Z.; Yao, B.; Wang, S.; Fan, X.; Wu, G.; Yang, H.; Yin, H.; Yang, S.; Harris, R.C. Intrarenal dopamine deficiency leads to hypertension and decreased longevity in mice. J. Clin. Investig. 2011, 121, 2845–2854. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Jose, P.A. Dopamine receptors: Important antihypertensive counterbalance against hypertensive factors. Hypertension 2011, 57, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Carey, R.M. Theodore Cooper Lecture: Renal dopamine system: Paracrine regulator of sodium homeostasis and blood pressure. Hypertension 2001, 38, 297–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haney, M.; Ward, A.S.; Foltin, R.W.; Fischman, M.W. Effects of ecopipam, a selective dopamine D1 antagonist, on smoked cocaine self-administration by humans. Psychopharmacology 2001, 155, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Henderson, D.C.; Daley, T.B.; Kunkel, L.; Rodrigues-Scott, M.; Koul, P.; Hayden, D. Clozapine and hypertension: A chart review of 82 patients. J. Clin. Psychiatry 2004, 65, 686–689. [Google Scholar] [CrossRef] [PubMed]
- Armando, I.; Konkalmatt, P.; Felder, R.A.; Jose, P.A. The renal dopaminergic system: Novel diagnostic and therapeutic approaches in hypertension and kidney disease. Transl. Res. 2015, 165, 505–511. [Google Scholar] [CrossRef] [Green Version]
- Gildea, J.J.; Xu, P.; Carlson, J.M.; Gaglione, R.T.; Bigler Wang, D.; Kemp, B.A.; Reyes, C.M.; McGrath, H.E.; Carey, R.M.; Jose, P.A.; et al. The sodium-bicarbonate cotransporter NBCe2 (slc4a5) expressed in human renal proximal tubules shows increased apical expression under high-salt conditions. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 309, R1447–R1459. [Google Scholar] [CrossRef] [Green Version]
- Kunimi, M.; Seki, G.; Hara, C.; Taniguchi, S.; Uwatoko, S.; Goto, A.; Kimura, S.; Fujita, T. Dopamine inhibits renal Na+:HCO3- cotransporter in rabbits and normotensive rats but not in spontaneously hypertensive rats. Kidney Int. 2000, 57, 534–543. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.Q.; Felder, R.A.; Carey, R.M. Selective inhibition of the renal dopamine subtype D1A receptor induces antinatriuresis in conscious rats. Hypertension 1999, 33, 504–510. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, F.E.; Drago, J.; Felder, R.A.; Printz, M.P.; Eisner, G.M.; Robillard, J.E.; Sibley, D.R.; Westphal, H.J.; Jose, P.A. Role of the D1A dopamine receptor in the pathogenesis of genetic hypertension. J. Clin. Investig. 1996, 97, 2283–2288. [Google Scholar] [CrossRef]
- Bacic, D.; Kaissling, B.; McLeroy, P.; Zou, L.; Baum, M.; Moe, O.W. Dopamine acutely decreases apical membrane Na/H exchanger NHE3 protein in mouse renal proximal tubule. Kidney Int. 2003, 64, 2133–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felder, C.C.; Albrecht, F.E.; Campbell, T.; Eisner, G.M.; Jose, P.A. cAMP-independent, G protein-linked inhibition of Na+/H+ exchange in renal brush border by D1 dopamine agonists. Am. J. Physiol. 1993, 264, F1032–F1037. [Google Scholar] [CrossRef] [PubMed]
- Gesek, F.A.; Schoolwerth, A.C. Hormonal interactions with the proximal Na(+)-H+ exchanger. Am. J. Physiol. 1990, 258, F514–F521. [Google Scholar] [CrossRef]
- Gesek, F.A.; Schoolwerth, A.C. Hormone responses of proximal Na(+)-H+ exchanger in spontaneously hypertensive rats. Am. J. Physiol. 1991, 261, F526–F536. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.C.; Fan, L.; Crowder, L.A.; Karim-Jimenez, Z.; Murer, H.; Moe, O.W. Dopamine acutely stimulates Na+/H+ exchanger (NHE3) endocytosis via clathrin-coated vesicles: Dependence on protein kinase A-mediated NHE3 phosphorylation. J. Biol. Chem. 2001, 276, 26906–26915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Li, W.; Du, P.; Yu, K.Q.; Fu, W. Molecular insights into the D1R agonist and D2R/D3R antagonist effects of the natural product (-)-stepholidine: Molecular modeling and dynamics simulations. J. Phys. Chem. B 2012, 116, 8121–8130. [Google Scholar] [CrossRef] [PubMed]
- Mohr, P.; Decker, M.; Enzensperger, C.; Lehmann, J. Dopamine/serotonin receptor ligands. 12(1): SAR studies on hexahydro-dibenz[d,g]azecines lead to 4-chloro-7-methyl-5,6,7,8,9,14-hexahydrodibenz[d,g]azecin-3-ol, the first picomolar D5-selective dopamine-receptor antagonist. J. Med. Chem. 2006, 49, 2110–2116. [Google Scholar] [CrossRef]
- Gildea, J.J.; Shah, I.T.; Van Sciver, R.E.; Israel, J.A.; Enzensperger, C.; McGrath, H.E.; Jose, P.A.; Felder, R.A. The cooperative roles of the dopamine receptors, D1R and D5R, on the regulation of renal sodium transport. Kidney Int. 2014, 86, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Narkar, V.; Hussain, T.; Lokhandwala, M. Role of tyrosine kinase and p44/42 MAPK in D(2)-like receptor-mediated stimulation of Na(+), K(+)-ATPase in kidney. Am. J. Physiol. Ren. Physiol. 2002, 282, F697–F702. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, I.; Walk, S.F.; Jose, P.A.; Felder, R.A. Dopamine D2L receptors stimulate Na+/K(+)-ATPase activity in murine LTK- cells. Mol. Pharmacol. 1996, 49, 373–378. [Google Scholar]
- Zhang, Y.; Fu, C.; Asico, L.D.; Villar, V.A.; Ren, H.; He, D.; Wang, Z.; Yang, J.; Jose, P.A.; Zeng, C. Role of Gα(12)- and Gα(13)-protein subunit linkage of D(3) dopamine receptors in the natriuretic effect of D(3) dopamine receptor in kidney. Hypertens. Res. 2011, 34, 1011–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Ren, H.; Lu, X.; He, D.; Han, Y.; Wang, H.; Zeng, C.; Shi, W. Inhibition of D4 Dopamine Receptors on Insulin Receptor Expression and Effect in Renal Proximal Tubule Cells. J. Am. Heart Assoc. 2016, 5, e002448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, A.J.; Meijer, S.; Wesseling, H.; Donker, A.J.; Reitsma, W.D. Dissociation of renal vasodilator and natriuretic effects of dopamine during sulpiride infusion in normal man. Eur. J. Clin. Pharmacol. 1990, 39, 221–226. [Google Scholar] [CrossRef]
- Luippold, G.; Zimmermann, C.; Mai, M.; Kloor, D.; Starck, D.; Gross, G.; Mühlbauer, B. Dopamine D(3) receptors and salt-dependent hypertension. J. Am. Soc. Nephrol. 2001, 12, 2272–2279. [Google Scholar] [PubMed]
- Fantini, J.; Di Scala, C.; Baier, C.J.; Barrantes, F.J. Molecular mechanisms of protein-cholesterol interactions in plasma membranes: Functional distinction between topological (tilted) and consensus (CARC/CRAC) domains. Chem. Phys. Lipids 2016, 199, 52–60. [Google Scholar] [CrossRef]
- Chidlow, J.H.; Sessa, W.C. Caveolae, caveolins, and cavins: Complex control of cellular signalling and inflammation. Cardiovasc. Res. 2010, 86, 219–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, P.; Sun, M.; Villar, V.A.; Zhang, Y.; Weinman, E.J.; Felder, R.A.; Jose, P.A. Differential dopamine receptor subtype regulation of adenylyl cyclases in lipid rafts in human embryonic kidney and renal proximal tubule cells. Cell. Signal. 2014, 26, 2521–2529. [Google Scholar] [CrossRef] [Green Version]
- Han, W.; Li, H.; Villar, V.A.; Pascua, A.M.; Dajani, M.I.; Wang, X.; Natarajan, A.; Quinn, M.T.; Felder, R.A.; Jose, P.A.; et al. Lipid rafts keep NADPH oxidase in the inactive state in human renal proximal tubule cells. Hypertension 2008, 51, 481–487. [Google Scholar] [CrossRef] [Green Version]
- Villar, V.A.; Armando, I.; Sanada, H.; Frazer, L.C.; Russo, C.M.; Notario, P.M.; Lee, H.; Comisky, L.; Russell, H.A.; Yang, Y.; et al. Novel role of sorting nexin 5 in renal D(1) dopamine receptor trafficking and function: Implications for hypertension. FASEB J. 2013, 27, 1808–1819. [Google Scholar] [CrossRef] [Green Version]
- Tiu, A.C.; Yang, J.; Asico, L.D.; Konkalmatt, P.; Zheng, X.; Cuevas, S.; Wang, X.; Lee, H.; Mazhar, M.; Felder, R.A.; et al. Lipid rafts are required for effective renal D1 dopamine receptor function. FASEB J. 2020, 34, 6999–7017. [Google Scholar] [CrossRef] [Green Version]
- Gildea, J.J.; Israel, J.A.; Johnson, A.K.; Zhang, J.; Jose, P.A.; Felder, R.A. Caveolin-1 and dopamine-mediated internalization of NaKATPase in human renal proximal tubule cells. Hypertension 2009, 54, 1070–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gildea, J.J.; Kemp, B.A.; Howell, N.L.; Van Sciver, R.E.; Carey, R.M.; Felder, R.A. Inhibition of renal caveolin-1 reduces natriuresis and produces hypertension in sodium-loaded rats. Am. J. Physiol. Ren. Physiol. 2011, 300, F914–F920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, M.M.; Hasbi, A.; Mattocks, M.; Fan, T.; O’Dowd, B.F.; George, S.R. Regulation of D1 dopamine receptor trafficking and signaling by caveolin-1. Mol. Pharmacol. 2007, 72, 1157–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somkuwar, S.S.; Fannon, M.J.; Head, B.P.; Mandyam, C.D. Methamphetamine reduces expression of caveolin-1 in the dorsal striatum: Implication for dysregulation of neuronal function. Neuroscience 2016, 328, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Niu, M.; Zhou, Z.; Zheng, X.; Zhang, L.; Tian, Y.; Yu, X.; Bu, G.; Xu, H.; Ma, Q.; et al. VPS35 regulates cell surface recycling and signaling of dopamine receptor D1. Neurobiol. Aging 2016, 46, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Yu, P.; Yang, Z.; Jones, J.E.; Wang, Z.; Owens, S.A.; Mueller, S.C.; Felder, R.A.; Jose, P.A. D1 dopamine receptor signaling involves caveolin-2 in HEK-293 cells. Kidney Int. 2004, 66, 2167–2180. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Xi, B. Pooled analyses of the associations of polymorphisms in the GRK4 and EMILIN1 genes with hypertension risk. Int. J. Med. Sci. 2012, 9, 274–279. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Villar, V.A.; Jones, J.E.; Jose, P.A.; Zeng, C. G protein-coupled receptor kinase 4: Role in hypertension. Hypertension 2015, 65, 1148–1155. [Google Scholar] [CrossRef]
- Felder, R.A.; Sanada, H.; Xu, J.; Yu, P.Y.; Wang, Z.; Watanabe, H.; Asico, L.D.; Wang, W.; Zheng, S.; Yamaguchi, I.; et al. G protein-coupled receptor kinase 4 gene variants in human essential hypertension. Proc. Natl. Acad. Sci. USA 2002, 99, 3872–3877. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Zeng, C.; Villar, V.A.; Chen, S.Y.; Konkalmatt, P.; Wang, X.; Asico, L.D.; Jones, J.E.; Yang, Y.; Sanada, H.; et al. Human GRK4γ142V Variant Promotes Angiotensin II Type I Receptor-Mediated Hypertension via Renal Histone Deacetylase Type 1 Inhibition. Hypertension 2016, 67, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Allen, S.J.; Parthasarathy, G.; Darke, P.L.; Diehl, R.E.; Ford, R.E.; Hall, D.L.; Johnson, S.A.; Reid, J.C.; Rickert, K.W.; Shipman, J.M.; et al. Structure and Function of the Hypertension Variant A486V of G Protein-coupled Receptor Kinase 4. J. Biol. Chem. 2015, 290, 20360–20373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Villar, V.A.; Armando, I.; Jose, P.A.; Zeng, C. G Protein-Coupled Receptor Kinases: Crucial Regulators of Blood Pressure. J. Am. Heart Assoc. 2016, 5, e003519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rayner, B.; Ramesar, R. The importance of G protein-coupled receptor kinase 4 (GRK4) in pathogenesis of salt sensitivity, salt sensitive hypertension and response to antihypertensive treatment. Int. J. Mol. Sci. 2015, 16, 5741–5749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, H.; Xu, J.; Bengra, C.; Jose, P.A.; Felder, R.A. Desensitization of human renal D1 dopamine receptors by G protein-coupled receptor kinase 4. Kidney Int. 2002, 62, 790–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanada, H.; Yoneda, M.; Yatabe, J.; Williams, S.M.; Bartlett, J.; White, M.J.; Gordon, L.N.; Felder, R.A.; Eisner, G.M.; Armando, I.; et al. Common variants of the G protein-coupled receptor type 4 are associated with human essential hypertension and predict the blood pressure response to angiotensin receptor blockade. Pharm. J. 2016, 16, 3–9. [Google Scholar] [CrossRef]
- Kimura, L.; Angeli, C.B.; Auricchio, M.T.; Fernandes, G.R.; Pereira, A.C.; Vicente, J.P.; Pereira, T.V.; Mingroni-Netto, R.C. Multilocus family-based association analysis of seven candidate polymorphisms with essential hypertension in an African-derived semi-isolated Brazilian population. Int. J. Hypertens. 2012, 2012, 859219. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Han, W.; Villar, V.A.; Keever, L.B.; Lu, Q.; Hopfer, U.; Quinn, M.T.; Felder, R.A.; Jose, P.A.; Yu, P. D1-like receptors regulate NADPH oxidase activity and subunit expression in lipid raft microdomains of renal proximal tubule cells. Hypertension 2009, 53, 1054–1061. [Google Scholar] [CrossRef] [Green Version]
- Hailstones, D.; Sleer, L.S.; Parton, R.G.; Stanley, K.K. Regulation of caveolin and caveolae by cholesterol in MDCK cells. J. Lipid Res. 1998, 39, 369–379. [Google Scholar]
- Wang, J.; Fedoseienko, A.; Chen, B.; Burstein, E.; Jia, D.; Billadeau, D.D. Endosomal receptor trafficking: Retromer and beyond. Traffic 2018, 19, 578–590. [Google Scholar] [CrossRef] [Green Version]
- Gallon, M.; Cullen, P.J. Retromer and sorting nexins in endosomal sorting. Biochem. Soc. Trans. 2015, 43, 33–47. [Google Scholar] [CrossRef]
- Teasdale, R.D.; Collins, B.M. Insights into the PX (phox-homology) domain and SNX (sorting nexin) protein families: Structures, functions and roles in disease. Biochem. J. 2012, 441, 39–59. [Google Scholar] [CrossRef] [PubMed]
- Bek, M.J.; Zheng, S.; Xu, J.; Yamaguchi, I.; Asico, L.D.; Sun, X.G.; Jose, P.A. Differential expression of adenylyl cyclases in the rat nephron. Kidney Int. 2001, 60, 890–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Yang, J.; Jones, J.E.; Villar, V.A.; Yu, P.; Armando, I.; Felder, R.A.; Jose, P.A. Sorting nexin 5 and dopamine d1 receptor regulate the expression of the insulin receptor in human renal proximal tubule cells. Endocrinology 2015, 156, 2211–2221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Villar, V.; Yang, J.A.; Jones, J.E.; Guo, Y.; Asico, L.D.; Armando, I.; Weinman, E.J.; Jose, P.A. Sorting nexin 19: A novel regulator of renal dopamine D1 receptor. Hypertension 2014, 64, A296. [Google Scholar]
- Heydorn, A.; Søndergaard, B.P.; Hadrup, N.; Holst, B.; Haft, C.R.; Schwartz, T.W. Distinct in vitro interaction pattern of dopamine receptor subtypes with adaptor proteins involved in post-endocytotic receptor targeting. FEBS Lett. 2004, 556, 276–280. [Google Scholar] [CrossRef] [Green Version]
- Villar, V.A.; Jones, J.E.; Armando, I.; Asico, L.D.; Escano, C.S.; Lee, H.; Wang, X.; Yang, Y.; Pascua-Crusan, A.M.; Palmes-Saloma, C.P.; et al. Sorting nexin 1 loss results in D5 dopamine receptor dysfunction in human renal proximal tubule cells and hypertension in mice. J. Biol. Chem. 2013, 288, 152–163. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Asico, L.D.; Beitelshees, A.L.; Feranil, J.B.; Wang, X.; Jones, J.E.; Armando, I.; Cuevas, S.G.; Schwartz, G.L.; Gums, J.G.; et al. Sorting nexin 1 loss results in increased oxidative stress and hypertension. FASEB J. 2020, 34, 7941–7957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.; Chen, C.; Guo, L.; Xu, Z.; Peng, X.; Wang, X.; Wang, J.; Wang, N.; Li, C.; Luo, X.; et al. Exposure to Maternal Diabetes Mellitus Causes Renal Dopamine D1 Receptor Dysfunction and Hypertension in Adult Rat Offspring. Hypertension 2018, 72, 962–970. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.R.; Kouyoumdzian, N.M.; Rukavina Mikusic, N.L.; Kravetz, M.C.; Rosón, M.I.; Rodríguez Fermepin, M.; Fernández, B.E. Renal dopaminergic system: Pathophysiological implications and clinical perspectives. World J. Nephrol. 2015, 4, 196–212. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Celver, J.; Octeau, J.C.; Kovoor, A. Plasma membrane compartmentalization of D2 dopamine receptors. J. Biol. Chem. 2013, 288, 12554–12568. [Google Scholar] [CrossRef] [Green Version]
- Genedani, S.; Guidolin, D.; Leo, G.; Filaferro, M.; Torvinen, M.; Woods, A.S.; Fuxe, K.; Ferré, S.; Agnati, L.F. Computer-assisted image analysis of caveolin-1 involvement in the internalization process of adenosine A2A-dopamine D2 receptor heterodimers. J. Mol. Neurosci. 2005, 26, 177–184. [Google Scholar] [CrossRef]
- Voulalas, P.J.; Schetz, J.; Undieh, A.S. Differential subcellular distribution of rat brain dopamine receptors and subtype-specific redistribution induced by cocaine. Mol. Cell. Neurosci. 2011, 46, 645–654. [Google Scholar] [CrossRef] [Green Version]
- Lan, H.; Teeter, M.M.; Gurevich, V.V.; Neve, K.A. An intracellular loop 2 amino acid residue determines differential binding of arrestin to the dopamine D2 and D3 receptors. Mol. Pharmacol. 2009, 75, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Wandelmer, J.; Dávalos, A.; de la Peña, G.; Cano, S.; Giera, M.; Canfrán-Duque, A.; Bracher, F.; Martín-Hidalgo, A.; Fernández-Hernando, C.; Lasunción, M.A.; et al. Haloperidol disrupts lipid rafts and impairs insulin signaling in SH-SY5Y cells. Neuroscience 2010, 167, 143–153. [Google Scholar] [CrossRef]
- Zhang, X.; Kim, K.M. Palmitoylation of the carboxyl-terminal tail of dopamine D4 receptor is required for surface expression, endocytosis, and signaling. Biochem. Biophys. Res. Commun. 2016, 479, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Free, R.B.; Namkung, Y.; Hazelwood, L.A.; Sibley, D.R. Sorting nexin-25 interacts with D1 and D2 dopamine receptors to regulate receptor expression and signaling. FASEB J. 2010, 21, 568–569. [Google Scholar] [CrossRef]
- Free, R.B.; Hazelwood, L.A.; Spalding, H.N.; Cabrera, D.M.; Sibley, D.R. Sorting nexin-25, a novel member of the dopamine receptor signalplex, up-regulates D1 and D2 dopamine receptor expression in HEK293 cells. FASEB J. 2007, 21, A423. [Google Scholar] [CrossRef]
- Rifkin, R.A.; Huyghe, D.; Li, X.; Parakala, M.; Aisenberg, E.; Moss, S.J.; Slesinger, P.A. GIRK currents in VTA dopamine neurons control the sensitivity of mice to cocaine-induced locomotor sensitization. Proc. Natl. Acad. Sci. USA 2018, 115, E9479–E9488. [Google Scholar] [CrossRef] [Green Version]
- Mystek, P.; Dutka, P.; Tworzydło, M.; Dziedzicka-Wasylewska, M.; Polit, A. The role of cholesterol and sphingolipids in the dopamine D1 receptor and G protein distribution in the plasma membrane. Biochem. Biophys. Acta 2016, 1861, 1775–1786. [Google Scholar]
Figure 1.
Cell membrane partitions into lipid raft and non-lipid raft microdomains.

Figure 2.
Caveola structure.

Figure 3.
Agonist-induced stimulation of G protein-coupled receptors (GPCRs) (e.g., dopamine receptor (1) and phosphorylation of GDP to GTP on the Gα subunit of the G protein (2)).
Figure 3.
Agonist-induced stimulation of G protein-coupled receptors (GPCRs) (e.g., dopamine receptor (1) and phosphorylation of GDP to GTP on the Gα subunit of the G protein (2)).

Figure 4.
Uncoupling of Gα subunit from Gβγ subunit, followed by activation GPCR kinases (GRKs), which phosphorylate amino acids in the third intracellular loop of the transmembrane segments and C-terminal tail (3a). The uncoupled Gα subunit activates downstream cellular (3b) and subsequent cellular response (3c).
Figure 4.
Uncoupling of Gα subunit from Gβγ subunit, followed by activation GPCR kinases (GRKs), which phosphorylate amino acids in the third intracellular loop of the transmembrane segments and C-terminal tail (3a). The uncoupled Gα subunit activates downstream cellular (3b) and subsequent cellular response (3c).

Figure 5.
The phosphorylation of amino acids in the third intracellular loop of the transmembrane segments and C-terminal tail leads to the activation of β-arrestins (5a).
Figure 5.
The phosphorylation of amino acids in the third intracellular loop of the transmembrane segments and C-terminal tail leads to the activation of β-arrestins (5a).

Figure 6.
Dynamin and clathrin associate with the GPCR (dopamine receptor) and mediate its endocytosis by means of the adaptor protein 2 (AP2) (6a).
Figure 6.
Dynamin and clathrin associate with the GPCR (dopamine receptor) and mediate its endocytosis by means of the adaptor protein 2 (AP2) (6a).

Figure 7.
Endocytosed GPCR (dopamine receptor) is packaged in an early endosome (7a).

Figure 8.
The endocytosed GPCR (dopamine receptor) binds to sorting nexin (SNX) that sends the GPCR either to a recycling endosome or lysosome (8a). Receptors targeted to lysosomes are degraded.
Figure 8.
The endocytosed GPCR (dopamine receptor) binds to sorting nexin (SNX) that sends the GPCR either to a recycling endosome or lysosome (8a). Receptors targeted to lysosomes are degraded.

Figure 9.
The resensitized GPCR (dopamine receptor) is targeted to the lipid raft microdomain by SNX (9a) and is ready to bind to a new agonist (9a).
Figure 9.
The resensitized GPCR (dopamine receptor) is targeted to the lipid raft microdomain by SNX (9a) and is ready to bind to a new agonist (9a).

Figure 10.
Structural differences between D1-like receptor (left side of picture) and D2-like receptors (right side of picture).
Figure 10.
Structural differences between D1-like receptor (left side of picture) and D2-like receptors (right side of picture).

Figure 11.
D1-like receptors are linked to Gαs subunit.

Figure 12.
D1-like receptors are linked to Gαq subunit.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Transmembrane segments of D1-like and D2-like dopamine receptors.
| D1R and D5R | D2R and D3R | D2R and D4R | |
|---|---|---|---|
| Amino acid sequence conservation within transmembrane domain | 80% | 75% | 53% |
Table 2.
NH2 terminal tail in D1-like and D2-like receptors.
| D1R and D5R | D2R | D3R | D4R | |
|---|---|---|---|---|
| Number of consensus N-glycosylation sites | 2 | 4 | 3 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Martinez, V.J.; Asico, L.D.; Jose, P.A.; Tiu, A.C. Lipid Rafts and Dopamine Receptor Signaling. Int. J. Mol. Sci. 2020, 21, 8909. https://doi.org/10.3390/ijms21238909
AMA Style
Martinez VJ, Asico LD, Jose PA, Tiu AC. Lipid Rafts and Dopamine Receptor Signaling. International Journal of Molecular Sciences. 2020; 21(23):8909. https://doi.org/10.3390/ijms21238909
Chicago/Turabian StyleMartinez, Victor J., Laureano D. Asico, Pedro A. Jose, and Andrew C. Tiu. 2020. "Lipid Rafts and Dopamine Receptor Signaling" International Journal of Molecular Sciences 21, no. 23: 8909. https://doi.org/10.3390/ijms21238909
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.