Effects of Hydrogen Sulfide Donor NaHS on Porcine Vascular Wall-Mesenchymal Stem Cells

 , ,
, ,  , , and
, , and 
Abstract
:1. Introduction
2. Results
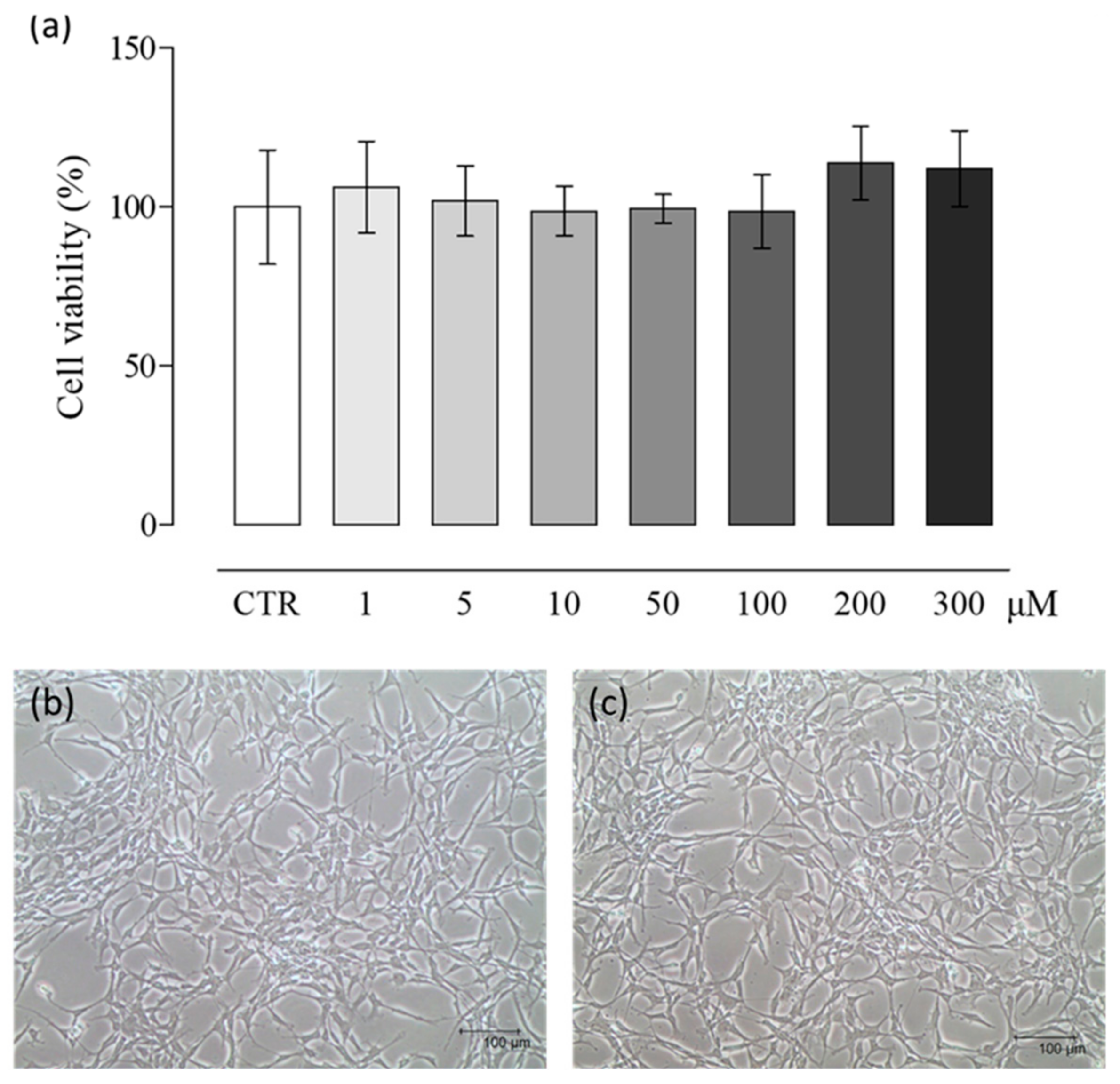
2.1. NaHS Effects on pVW–MSC Viability
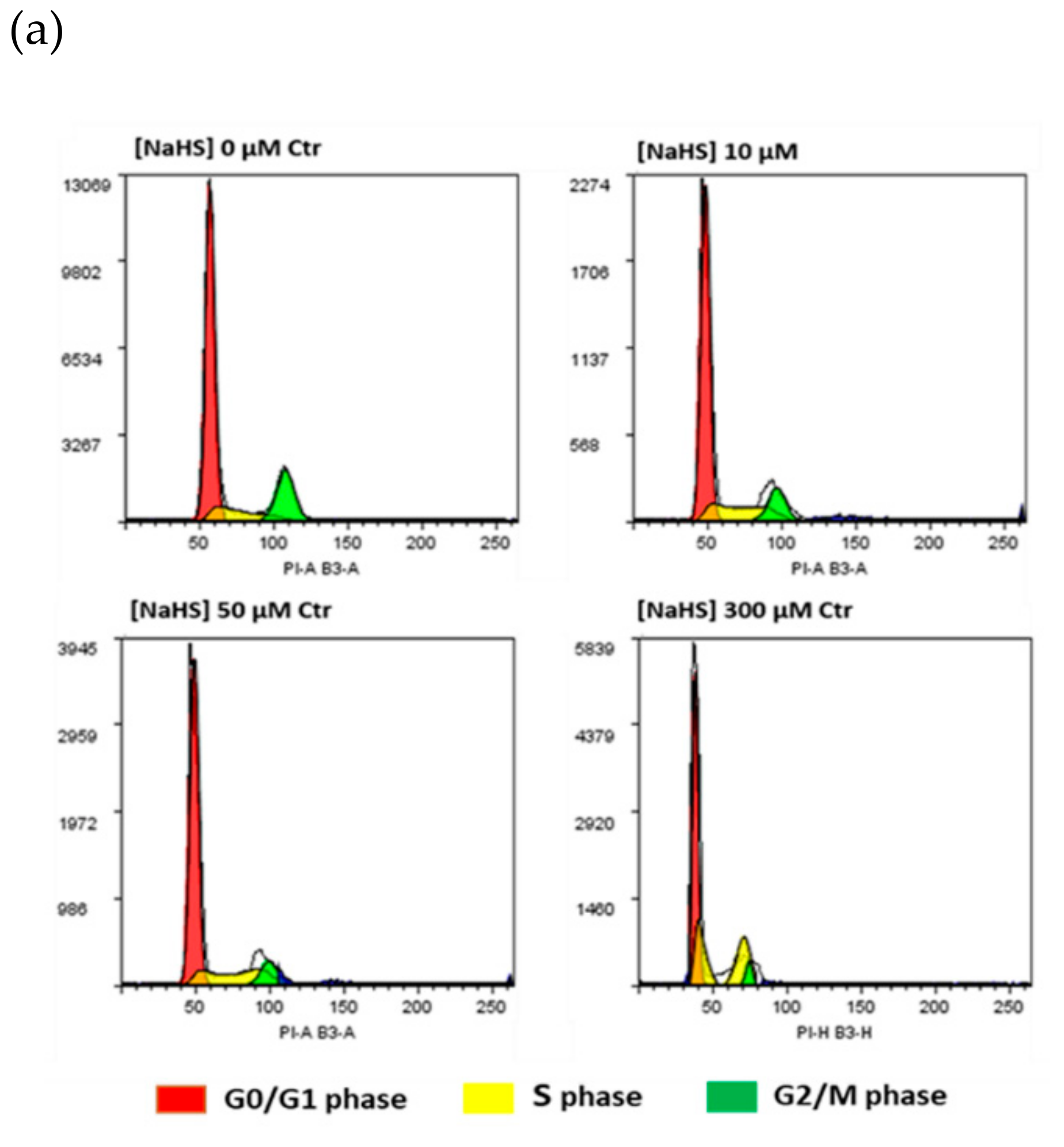
2.2. NaHS Effects on pVW–MSC Cell Cycle and ROS Production
2.3. NaHS Effects on pVW–MSC Mesenchymal Gene Expression and Angiogenic Property
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Cultures
4.3. Cell Viability
4.4. Cell Cycle Analysis
4.5. Reactive Oxygen Species Evaluation
4.6. Gene Expression Analysis after Long Term Culture with NaHS
4.7. In Vitro Angiogenesis Spheroid Assay
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| H2S | Hydrogen sulfide |
| VW–MSCs | Vascular wall–mesenchymal stem cells |
| CD90 | THYmocyte differentiation antigen 1 |
| NG2 | Neural/glial antigen 2 |
| PDGFR-β | Platelet-derived growth factor receptor beta |
| α-SMA | Alpha-smooth muscle actine |
| pVW–MSCs | Porcine vascular wall–mesenchymal stem cells |
| CD31 | Platelet endothelial cell adhesion molecule |
| NO | Nitric oxide |
| CO | Carbon monoxide |
| ROS | Reactive oxygen species |
| VSCs | Vascular stem cells |
References
- Zhao, Y.; Biggs, T.D.; Xian, M. Hydrogen sulfide (H2S) releasing agents: Chemistry and biological applications. Chem. Commun. 2014, 50, 11788–11805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, K.; Kimura, H. The possible role of hydrogen sulfide as an endogenous neuromodulator. J. Neurosci. 1996, 16, 1066–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, K.R. A practical look at the chemistry and biology of hydrogen sulfide. Antioxid. Redox Signal. 2012, 17, 32–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandiver, M.S.; Snyder, S.H. Hydrogen sulfide: A gasotransmitter of clinical relevance. J. Mol. Med. 2012, 90, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Powell, C.R.; Dillon, K.M.; Matson, J.B. A review of hydrogen sulfide (H2S) donors: Chemistry and potential therapeutic applications. Biochem. Pharmacol. 2018, 149, 110–123. [Google Scholar] [CrossRef]
- Martelli, A.; Testai, L.; Breschi, M.C.; Blandizzi, C.; Virdis, A.; Taddei, S.; Calderone, V. Hydrogen sulphide: Novel opportunity for drug discovery. Med. Res. Rev. 2012, 32, 1093–1130. [Google Scholar] [CrossRef]
- Bełtowski, J. Hydrogen sulfide in pharmacology and medicine—An update. Pharmacol. Rep. 2015, 67, 647–658. [Google Scholar] [CrossRef]
- Caliendo, G.; Cirino, G.; Santagada, V.; Wallace, J.L. Synthesis and Biological Effects of Hydrogen Sulfide (H2S): Development of H2S-Releasing Drugs as Pharmaceuticals. J. Med. Chem. 2010, 53, 6275–6286. [Google Scholar] [CrossRef]
- Deplancke, B.; Gaskins, H.R. Hydrogen sulfide induces serum-independent cell cycle entry in nontransformed rat intestinal epithelial cells. FASEB J. 2003, 17, 1310–1312. [Google Scholar] [CrossRef] [Green Version]
- Baskar, R.; Li, L.; Moore, P.K. Hydrogen sulfide-induces DNA damage and changes in apoptotic gene expression in human lung fibroblast cells. FASEB J. 2007, 21, 247–255. [Google Scholar] [CrossRef]
- Xiao, A.Y.; Maynard, M.R.; Piett, C.G.; Nagel, Z.D.; Alexander, J.S.; Kevil, C.G.; Berridge, M.V.; Pattillo, C.B.; Rosen, L.R.; Miriyala, S.; et al. Sodium sulfide selectively induces oxidative stress, DNA damage, and mitochondrial dysfunction and radiosensitizes glioblastoma (GBM) cells. Redox Biol 2019, 26, 101220. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, M.; Rajapakse, A.; Shen, X.; Gates, K.S. Generation of DNA-damaging reactive oxygen species via the autoxidation of hydrogen sulfide under physiologically relevant conditions: Chemistry relevant to both the genotoxic and cell signaling properties of H(2)S. Chem. Res. Toxicol. 2012, 25, 1609–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, K.; Ju, Y.; Li, S.; Altaany, Z.; Wang, R.; Yang, G. S-sulfhydration of MEK1 leads to PARP-1 activation and DNA damage repair. EMBO Rep. 2014, 15, 792–800. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Wang, Z.; Zhan, J.; Zhang, Q.; Wang, J.; Zhang, Q.; Xian, X.; Luan, Q.; Hao, A. Hydrogen sulfide promotes proliferation and neuronal differentiation of neural stem cells and protects hypoxia-induced decrease in hippocampal neurogenesis. Pharmacol. Biochem. Behav. 2014, 116, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wei, H.; Kong, G.; Shim, W.; Zhang, G. Hydrogen sulfide augments the proliferation and survival of human induced pluripotent stem cell-derived mesenchymal stromal cells through inhibition of BKCa. Cytotherapy 2013, 15, 1395–1405. [Google Scholar] [CrossRef] [PubMed]
- Pu, H.; Hua, Y. Hydrogen sulfide regulates bone remodeling and promotes orthodontic tooth movement. Mol. Med. Rep. 2017, 16, 9415–9422. [Google Scholar] [CrossRef]
- Yang, R.; Liu, Y.; Yu, T.; Liu, D.; Shi, S.; Zhou, Y.; Zhou, Y. Hydrogen sulfide maintains dental pulp stem cell function via TRPV1-mediated calcium influx. Cell Death Discov. 2018, 4, 1. [Google Scholar] [CrossRef]
- Aykan, A.; Ozturk, S.; Sahin, I.; Avcu, F.; Sagkan, R.I.; Isik, S. The Effects of Hydrogen Sulfide on Adipocyte Viability in Human Adipocyte and Adipocyte-Derived Mesenchymal Stem Cell Cultures Under Ischemic Conditions. Ann. Plast. Surg. 2015, 75, 657–665. [Google Scholar] [CrossRef]
- Guo, Z.; Li, C.S.; Wang, C.M.; Xie, Y.J.; Wang, A.L. CSE/H2S system protects mesenchymal stem cells from hypoxia and serum deprivation-induced apoptosis via mitochondrial injury, endoplasmic reticulum stress and PI3K/Akt activation pathways. Mol. Med. Rep. 2015, 12, 2128–2134. [Google Scholar] [CrossRef]
- Xie, X.; Sun, A.; Zhu, W.; Huang, Z.; Hu, X.; Jia, J.; Zou, Y.; Ge, J. Transplantation of mesenchymal stem cells preconditioned with hydrogen sulfide enhances repair of myocardial infarction in rats. Tohoku J. Exp. Med. 2012, 226, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.C.; Sohn, E.-H.; Lee, S.R. Hydrogen Sulfide as a Potential Alternative for the Treatment of Myocardial Fibrosis. Oxidative Med. Cell. Longev. 2020, 2020, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Psaltis, P.J.; Simari, R.D. Vascular wall progenitor cells in health and disease. Circ. Res. 2015, 116, 1392–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Invernici, G.; Emanueli, C.; Madeddu, P.; Cristini, S.; Gadau, S.; Benetti, A.; Ciusani, E.; Stassi, G.; Siragusa, M.; Nicosia, R.; et al. Human fetal aorta contains vascular progenitor cells capable of inducing vasculogenesis, angiogenesis, and myogenesis in vitro and in a murine model of peripheral ischemia. Am. J. Pathol. 2007, 170, 1879–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crisan, M.; Yap, S.; Casteilla, L.; Chen, C.-W.; Corselli, M.; Park, T.S.; Andriolo, G.; Sun, B.; Zheng, B.; Zhang, L.; et al. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell 2008, 3, 301–313. [Google Scholar] [CrossRef] [Green Version]
- Pasquinelli, G.; Pacilli, A.; Alviano, F.; Foroni, L.; Ricci, F.; Valente, S.; Orrico, C.; Lanzoni, G.; Buzzi, M.; Luigi-Tazzari, P.; et al. Multidistrict human mesenchymal vascular cells: Pluripotency and stemness characteristics. Cytotherapy 2010, 12, 275–287. [Google Scholar] [CrossRef]
- Zaniboni, A.; Bernardini, C.; Bertocchi, M.; Zannoni, A.; Bianchi, F.; Avallone, G.; Mangano, C.; Sarli, G.; Calzà, L.; Bacci, M.L.; et al. In vitro differentiation of porcine aortic vascular precursor cells to endothelial and vascular smooth muscle cells. Am. J. Physiol. Cell Physiol. 2015, 309, C320–C331. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.; Li, X. PDGFs and their receptors in vascular stem/progenitor cells: Functions and therapeutic potential in retinal vasculopathy. Mol. Asp. Med. 2018, 62, 22–32. [Google Scholar] [CrossRef]
- Mathew, S.A.; Naik, C.; Cahill, P.A.; Bhonde, R.R. Placental mesenchymal stromal cells as an alternative tool for therapeutic angiogenesis. Cell. Mol. Life Sci. 2020, 77, 253–265. [Google Scholar] [CrossRef]
- Zaniboni, A.; Bernardini, C.; Alessandri, M.; Mangano, C.; Zannoni, A.; Bianchi, F.; Sarli, G.; Calzà, L.; Bacci, M.L.; Forni, M. Cells derived from porcine aorta tunica media show mesenchymal stromal-like cell properties in in vitro culture. Am. J. Physiol. Cell Physiol. 2014, 306, C322–C333. [Google Scholar] [CrossRef] [Green Version]
- Bernardini, C.; Bertocchi, M.; Zannoni, A.; Salaroli, R.; Tubon, I.; Dothel, G.; Fernandez, M.; Bacci, M.L.; Calzà, L.; Forni, M. Constitutive and LPS-stimulated secretome of porcine Vascular Wall-Mesenchymal Stem Cells exerts effects on in vitro endothelial angiogenesis. BMC Vet. Res. 2019, 15. [Google Scholar] [CrossRef]
- Dothel, G.; Bernardini, C.; Zannoni, A.; Spirito, M.R.; Salaroli, R.; Bacci, M.L.; Forni, M.; Ponti, F.D. Ex vivo effect of vascular wall stromal cells secretome on enteric ganglia. World J. Gastroenterol. 2019, 25, 4892–4903. [Google Scholar] [CrossRef]
- Attene-Ramos, M.S.; Wagner, E.D.; Plewa, M.J.; Gaskins, H.R. Evidence that hydrogen sulfide is a genotoxic agent. Mol. Cancer Res. 2006, 4, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Nesci, S.; Bernardini, C.; Salaroli, R.; Zannoni, A.; Trombetti, F.; Ventrella, V.; Pagliarani, A.; Forni, M. Characterization of metabolic profiles and lipopolysaccharide effects on porcine vascular wall mesenchymal stem cells. J. Cell. Physiol. 2019. [Google Scholar] [CrossRef]
- Andrae, J.; Gallini, R.; Betsholtz, C. Role of platelet-derived growth factors in physiology and medicine. Genes Dev. 2008, 22, 1276–1312. [Google Scholar] [CrossRef] [Green Version]
- Brekke, C.; Lundervold, A.; Enger, P.Ø.; Brekken, C.; Stålsett, E.; Pedersen, T.B.; Haraldseth, O.; Krüger, P.G.; Bjerkvig, R.; Chekenya, M. NG2 expression regulates vascular morphology and function in human brain tumours. Neuroimage 2006, 29, 965–976. [Google Scholar] [CrossRef]
- Xie, L.; Zeng, X.; Hu, J.; Chen, Q. Characterization of Nestin, a Selective Marker for Bone Marrow Derived Mesenchymal Stem Cells. Stem Cells Int. 2015, 2015, 762098. [Google Scholar] [CrossRef] [Green Version]
- Zahra, F.T.; Choleva, E.; Sajib, M.S.; Papadimitriou, E.; Mikelis, C.M. In Vitro Spheroid Sprouting Assay of Angiogenesis. In The Extracellular Matrix: Methods and Protocols; Methods in Molecular Biology; Vigetti, D., Theocharis, A.D., Eds.; Springer: New York, NY, USA, 2019; ISBN 978-1-4939-9133-4. [Google Scholar]
- Sun, H.-J.; Wu, Z.-Y.; Nie, X.-W.; Bian, J.-S. Role of Endothelial Dysfunction in Cardiovascular Diseases: The Link Between Inflammation and Hydrogen Sulfide. Front. Pharmacol. 2019, 10, 1568. [Google Scholar] [CrossRef] [Green Version]
- Morbidelli, L.; Monti, M.; Terzuoli, E. Pharmacological Tools for the Study of H2S Contribution to Angiogenesis. Methods Mol. Biol. 2019, 2007, 151–166. [Google Scholar] [CrossRef]
- Cai, W.-J.; Wang, M.-J.; Moore, P.K.; Jin, H.-M.; Yao, T.; Zhu, Y.-C. The novel proangiogenic effect of hydrogen sulfide is dependent on Akt phosphorylation. Cardiovasc. Res. 2007, 76, 29–40. [Google Scholar] [CrossRef]
- Gambari, L.; Lisignoli, G.; Cattini, L.; Manferdini, C.; Facchini, A.; Grassi, F. Sodium hydrosulfide inhibits the differentiation of osteoclast progenitor cells via NRF2-dependent mechanism. Pharmacol. Res. 2014, 87, 99–112. [Google Scholar] [CrossRef]
- Sen, N. Functional and Molecular Insights of Hydrogen Sulfide Signaling and Protein Sulfhydration. J. Mol. Biol. 2017, 429, 543–561. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Rose, P.; Moore, P.K. Hydrogen sulfide and cell signaling. Annu. Rev. Pharmacol. Toxicol. 2011, 51, 169–187. [Google Scholar] [CrossRef] [Green Version]
- Fox, M.H. A model for the computer analysis of synchronous DNA distributions obtained by flow cytometry. Cytometry 1980, 1, 71–77. [Google Scholar] [CrossRef]
- Tubon, I.; Zannoni, A.; Bernardini, C.; Salaroli, R.; Bertocchi, M.; Mandrioli, R.; Vinueza, D.; Antognoni, F.; Forni, M. In Vitro Anti-Inflammatory Effect of Salvia sagittata Ethanolic Extract on Primary Cultures of Porcine Aortic Endothelial Cells. Oxid. Med. Cell. Longev. 2019, 2019. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sequence (5′–3′) | PCR Product (bp) | Accession Number | Reference |
|---|---|---|---|---|
| CD90 | For: GACTGCCGCCATGAGAATAC | 180 | NM_001146129.1 | Zaniboni et al., 2015 |
| Rev: GGTAGTGAAGCCTGATAAGTAGAG | ||||
| Nestin | For: CAGTGGTTCCAAGGCTTCTC | 163 | ENSSSCT00000027298 | Zaniboni et al., 2015 |
| Rev: CATAGGTGTGTCAAGGGTATCG | ||||
| NG2 | For: ACCACCTCCTCCTACAACTC | 104 | ENSSSCT00000002098 | Zaniboni et al., 2015 |
| Rev: GTCACTCAGCAGCATCTCTG | ||||
| PDGFRβ | For: GCAACGAGGTGGTCAACTTC | 111 | ENSSSCT00000015788 | Zaniboni et al., 2015 |
| Rev: GCAGGATAGAACGGATGTGG | ||||
| αSMA | For: CACGGCATCATCACCAACTG | 200 | NM_001164650 | Zaniboni et al., 2015 |
| Rev: ACCGCCTGAATAGCCACATAC | ||||
| GAPDH | For: ACATGGCCTCCAAGGAGTAAGA | 106 | NM_001206359 | Tubon et al., 2019 |
| Rev: GATCGAGTTGGGGCTGTGACT | ||||
| Probe: HEX-CCACCAACCCCAGCAAGAGCACGC-BHQ1 | ||||
| β-Actin | For: CTCGATCATGAAGTGCGACGT | 114 | KU672525.1 | Tubon et al., 2019 |
| Rev: GTGATCTCCTTCTGCATCCTGT | ||||
| Probe: FAM-ATCAGGAAGGACCTCTACGCCAACACGG-BHQ1 |
| Antibody | P. Number | Species | Supplier | Dilution |
|---|---|---|---|---|
| Anti CD31 | MCA1746 | Mouse | AbD Serotec | 1:50 |
| Anti Factor-VIII | RFF-8C/8 | Mouse | BioRad | 1:50 |
| Anti-Mouse-FITC | F4143 | Goat | Sigma-Aldrich | 1:200 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernardini, C.; La Mantia, D.; Nesci, S.; Salaroli, R.; Algieri, C.; Pagliarani, A.; Zannoni, A.; Forni, M. Effects of Hydrogen Sulfide Donor NaHS on Porcine Vascular Wall-Mesenchymal Stem Cells. Int. J. Mol. Sci. 2020, 21, 5267. https://doi.org/10.3390/ijms21155267
Bernardini C, La Mantia D, Nesci S, Salaroli R, Algieri C, Pagliarani A, Zannoni A, Forni M. Effects of Hydrogen Sulfide Donor NaHS on Porcine Vascular Wall-Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2020; 21(15):5267. https://doi.org/10.3390/ijms21155267
Chicago/Turabian StyleBernardini, Chiara, Debora La Mantia, Salvatore Nesci, Roberta Salaroli, Cristina Algieri, Alessandra Pagliarani, Augusta Zannoni, and Monica Forni. 2020. "Effects of Hydrogen Sulfide Donor NaHS on Porcine Vascular Wall-Mesenchymal Stem Cells" International Journal of Molecular Sciences 21, no. 15: 5267. https://doi.org/10.3390/ijms21155267