Neurexin Complexes
Overview
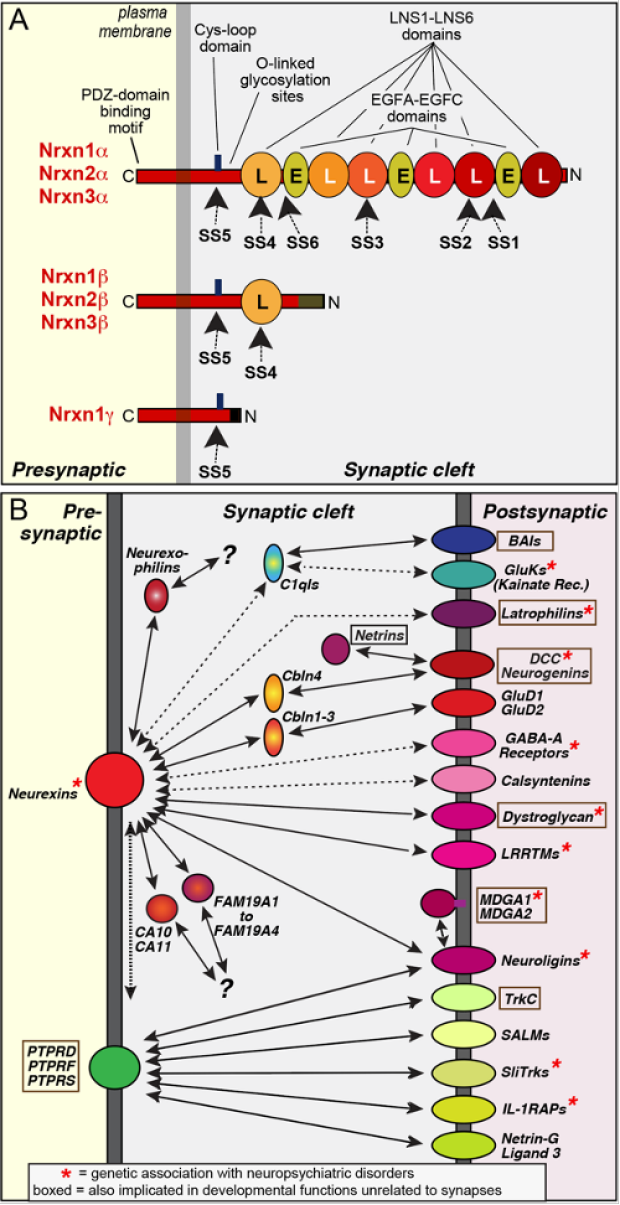
Neurexins are presynaptic adhesion molecules expressed from three genes (NRXN1-3 in humans, Nrxn1-3 in mice). All neurexin genes contain independent promoters for two principal forms (a- and b-neurexins), with b-neurexins being generally less abundant than a-neurexins1-3. In addition, the Nrxn1 gene includes a third promoter that drives expression of a g-form (see Figure panel A)4. Neurexin mRNAs are extensively alternatively spliced at 6 canonical sites (referred to as SS1-SS6), resulting in thousands of variants5-7. The expression of neurexins and their splice variants is temporally and spatially regulated in brain, conferring onto each neuron type a characteristic pattern of alternatively spliced neurexins5,8. Although all neurexins are highly homologous and include similar alternatively spliced sequences, their alternative splicing is not coordinately regulated but differs between Nrxn1, Nrxn2, and Nrxn3 in any given neuron8. Neurexins are, at least in part, localized to synapses where they form nanoclusters9. Moreover, Nrxn1-3 perform distinct functions that include the regulation of the presynaptic release probability, the recruitment of Ca2+-channels to presynaptic active zones, and the fine-tuning of postsynaptic receptor responses10. Neurexins are evolutionarily conserved3, with C. elegans containing a single neurexin gene (NRX-1) that encodes an a- and g- but not a b-isoform11. The alternative splicing of neurexins, however, appears to be absent from invertebrate neurexins. Finally, neurexins are closely related to the CNTNP family of adhesion molecules that exhibit a very similar domain structure12.
Neurexins interact with multiple extracellular ligands (see Figure panel B)10. Most of these ligands belong to gene families, resulting in >50 genes of potential ligands. Many neurexin-ligand interactions are regulated by neurexin alternative splicing (see details below). No canonical sequence motif is shared by neurexin ligands, suggesting diverse interaction surfaces. In addition to trans-synaptic neurexin ligands that are postsynaptic membrane proteins, neurexins interact with secreted ligands (such as CA10/11, FAM19’s [a.k.a. ‘TAFA’s’], and cerebellins). Finally, neurexin bind in cis to LAR-type phosphotyrosine receptor phosphatases (PTPRs) in an interaction that is possibly mediated via the post-translational heparan sulfate modification of neurexins13,14.
Neurexins likely interact with multiple ligands at the same time, although some ligands (e.g., LRRTMs and neuroligins, or dystroglycan and neurexophilins) compete with each other for binding. Different neurexins and different neurexin splice variants exhibit distinct affinities for various ligands, which in turn can also be subject to alternative splicing. Moreover, neurexin alternative splicing and neurexin ligand expression may be activity-dependent in some cases. Neurexins likely engage in a dynamic ‘fluid interaction network’ in which the precise synaptic complexes that are present at a time are determined by continuously changing concentrations and alternative splicing of various neurexins and their ligands. As a result, it is difficult to predict which neurexin-ligand complexes are present at a given synapse even if one knows which neurexin variants are expressed.
Knowledge gaps. More than 1,100 papers studying neurexins were published as of November 2023, neurexin-mutant mice and human neurons have been extensively studied, and atomic structures of neurexins and neurexin-ligand complexes are available. Yet, neurexins are poorly understood owing to their many variants, vast number of interactions, and diverse functions. Even fundamental questions are unaddressed. For example, how do neurexins perform some well-defined functions, such as their role in maintaining a normal presynaptic release probability and recruiting Ca2+-channels15-17, and what other functions do neurexins have? Do neurexins not only shape synapse properties but also help regulate synapse formation as indicated by the role of Nrxn2 in restricting synapse numbers18, and if so, by what mechanisms? How are neurexins coupled to active zones and what are their intracellular signaling mechanisms? Only three of the ten domains of neurexins (LNS2 and LNS6 domains and the ‘stalk’ region surrounding the Cys-loop domain), and only two of the six sites of alternative splicing (SS2 & SS4), have well-characterized ligand interactions – what do the other domains and the other sites of alternative splicing do, if anything? It is humbling to concede that even after decades of work by scores of labs, neurexins are still largely an enigma.
Synthesis. Given the large knowledge gaps, only tentative general conclusions are possible about neurexins. Nevertheless, an overall neurexin concept is beginning to emerge. Viewed together, current results reveal neurexins as signaling platforms with diverse functions mediated by multifarious trans-synaptic interactions. Via their many ligands, neurexins act as central control switches for synapse properties that may account for a large part of the functional diversity observed between different types of synapses.
References
1. Ushkaryov, Y.A., Petrenko, A.G., Geppert, M., and Südhof, T.C. (1992) Neurexins: Synaptic cell surface proteins related to the α-latrotoxin receptor and laminin. Science 257, 50-56.
2. Ushkaryov, Y.A., Hata, Y., Ichtchenko, K., Moomaw, C., Afendis, S., Slaughter, C.A., and Südhof, T.C. (1994) Conserved domain structure of ß-Neurexins. J. Biol. Chem. 269, 11987-11992.
3. Tabuchi, K., and Südhof, T.C. (2002) Structure and evolution of the neurexin genes: Insight into the mechanism of alternative splicing. Genomics 79, 849-859.
4. Sterky, F.H., Trotter, J.H., Lee, S., Recktenwald, C.V., Du, X., Zhou, B., Zhou, P., Schwenk, J., Fakler, B., and Südhof, T.C. (2017) The Carbonic Anhydrase-Related Protein CA10 Is An Evolutionarily Conserved Pan-Neurexin Ligand. Proc. Natl. Acad. Sci. U.S.A. 114, E1253-E1262.
5. Ullrich, B., Ushkaryov, Y.A., and Südhof, T.C. (1995) Cartography of neurexins: More than 1000 isoforms generated by alternative splicing and expressed in distinct subsets of neurons. Neuron 14, 497-507.
6. Treutlein, B., Gokce, O., Quake, S.R., and Südhof, T.C. (2014) Cartography of Neurexin Alternative Splicing Mapped by Single-Molecule Long-Read mRNA Sequencing. Proc. Natl. Acad. Sci. U.S.A. 111, E1291-1299.
7. Schreiner D, Nguyen TM, Russo G, Heber S, Patrignani A, Ahrné E, and Scheiffele, P. (2014) Targeted combinatorial alternative splicing generates brain region-specific repertoires of neurexins. Neuron 84, 386-398.
8. Fuccillo, M.V., Földy, C., Gökce, O., Rothwell, P.E., Sun, G., Malenka, R.C. and Südhof, T.C. (2015) Single-Cell mRNA Profiling Reveals Cell-Type Specific Expression of Neurexin Isoforms. Neuron 87, 326-340.
9. Trotter, J.H., Hao, J., Maxeiner, S., Tsetsenis, T., Liu, Z., Zhuang, X., and Südhof ,T.C. (2019) Synaptic Neurexin-1 Assembles into Dynamically Regulated Active Zone Nanoclusters. J. Cell Biology 218, 2677-2698.
10. Südhof, T.C. (2017) Synaptic Neurexin Complexes: A Molecular Code for the Logic of Neural Circuits. Cell 171, 745-769.
11. Kurshan PT, Merrill SA, Dong Y, Ding C, Hammarlund M, Bai J, Jorgensen EM, Shen K (2018) γ-Neurexin and Frizzled Mediate Parallel Synapse Assembly Pathways Antagonized by Receptor Endocytosis. Neuron 100, 150-166
12. Missler, M., Fernandez-Chacon, R., and Südhof, T.C. (1998) The making of neurexins. J. Neurochem. 71, 1339-1347.
13. Han KA, Kim YJ, Yoon TH, Kim H, Bae S, Um JW, Choi SY, and Ko J (2020) LAR-RPTPs Directly Interact with Neurexins to Coordinate Bidirectional Assembly of Molecular Machineries. J. Neurosci. 40, 8438-8462.
14. Roppongi RT, Dhume SH, Padmanabhan N, Silwal P, Zahra N, Karimi B, Bomkamp C, Patil CS, Champagne-Jorgensen K, Twilley RE, Zhang P, Jackson MF, and Siddiqui TJ (2020) LRRTMs Organize Synapses through Differential Engagement of Neurexin and PTPs. Neuron 106, 108-125.
15. Missler, M., Zhang, W., Rohlmann, A., Kattenstroth, G., Hammer, R.E., Gottmann, K., and Südhof, T.C. (2003) a-Neurexins Couple Ca2+-Channels to Synaptic Vesicle Exocytosis. Nature 423, 939-948.
16. Chen, L.Y., Jiang, M., Zhang, B., Gokce, O., and Südhof, T.C. (2017) Conditional Deletion of All Neurexins Defines Diversity of Essential Synaptic Organizer Functions for Neurexins. Neuron 94, 611-625.
17. Luo, F., Sclip, A., Jiang, M., and Südhof, T.C. (2020) Neurexins Cluster Ca2+ Channels within presynaptic Active Zone. EMBO J. 39, e103208.
18. Lin, P.Y., Chen, L.Y., Jiang, M., Trotter, J.H., Seigneur, E., and Südhof, T.C. (2023) Neurexin-2: An Inhibitory Neurexin That Restricts Excitatory Synapse Formation in the Hippocampus. Sci. Advances 9, eadd8856.
Neurexin-Neuroligin Complexes
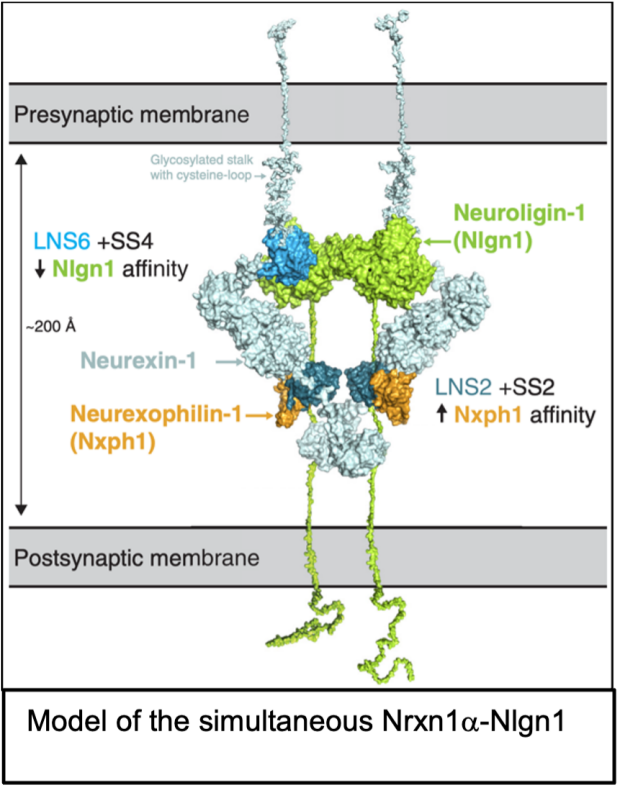
Properties. Neuroligins are type I membrane proteins composed of a large extracellular, constitutively dimeric esterase-like domain, a single transmembrane region, and a short cytoplasmic sequence (see figure)1-3. Mammals express four neuroligin genes (NLGN1-4 in humans and Nlgn1-4 in mice)2,3. Since the NLGN4 gene is located on the X and Y chromosomes and does not recombine, the slightly different NLGN4 alleles are also referred to as NLGN4X and NLGN4Y4.
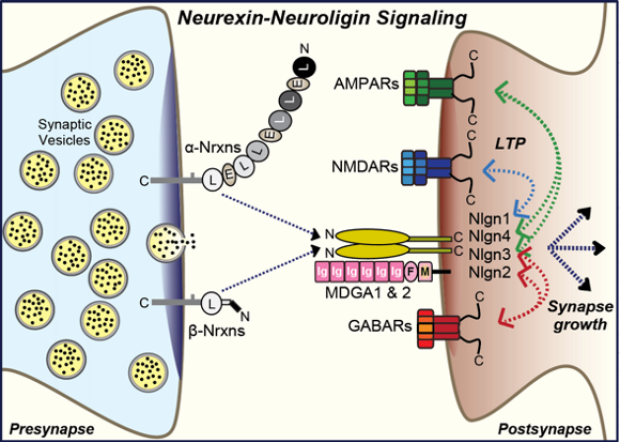
The four neuroligins exhibit distinct functional properties and localizations. Nlgn1 is specific for excitatory synapses, Nlgn2 is selectively localized to inhibitory and cholinergic synapses, Nlgn3 is present in both excitatory and inhibitory synapses, and Nlgn4 is found in glycinergic inhibitory synapses in rodents and in excitatory synapses in humans (reviewed in ref.5, 6). Mutations in neuroligin genes were reported in neuropsychiatric disorders, especially autism, with mutations in NLGN4X exhibiting the highest frequency7. Like neurexins, neuroligins are also expressed in glia but considerable controversy exists about their glial functions8.
Interactions. Neuroligins were discovered as neurexin ligands1 (hence their name), but neuroligins also bind to MDGA1 and MDGA2, which are postsynaptic Ig-domain proteins (see figure)9. Different neurexin/neuroligin and MDGA/neuroligin pairs exhibit distinct binding affinities, although these have only been partly characterized9-11. Neuroligins have two sites of alternative splicing (SSA and SSB), of which SSA is present in Nlgn1-3 but not in Nlgn4 and SSB in Nlgn1 but not in Nlgn2-410. We initially observed that Nlgn1 exclusively binds to neurexins lacking SS4 inserts (i.e., are SS4-)1,2, but later found that only Nlgn1 containing in insert in SSB (Nlgn1-SSB+, the predominant form in brain) is unable to bind to SS4+ neurexins, whereas Nlgn1-SSB- and all other neuroligins bind to both SS4+ and SS4- neurexins, albeit with different affinities10,11. A neuroligin cannot bind neurexins and MDGAs simultaneously, but it is plausible that in a neuroligin dimer one neuroligin subunit binds to a neurexin and the other neuroligin subunit to an MDGA. As a result, it is difficult to predict the state of Nrxn-Nlgn vs. Nlgn-MDGA complexes at a synapse, disregarding the added complexity provided by other ligands in shaping trans-synaptic neurexin complexes, such as cerebellins that have a higher affinity for SS4+ neurexins than neuroligins and that can displace neuroligins from neurexins12. In addition to the neurexin and MDGA interactions, Nlgn3 also binds to the N-terminal Ig-domains of the sigma isoform of phosphotyrosine receptor phosphatases (PTPRS)13, an interaction that may possibly also apply to other neuroligins.
Functions. Neuroligins perform major roles at synapses. As observed for neurexins, different neuroligins often have distinct, non-overlapping functions. In cultured mouse and/or human neurons, deletions of Nlgn1 selectively decrease AMPAR- and NMDAR-mediated synaptic transmission, deletions of Nlgn2 selectively decrease GABAR-mediated synaptic transmission, and deletions of Nlgn3 and Nlgn4 have little effects as single KOs, although they can aggravate some Nlgn1 or Nlgn2 deletion phenotypes in double KOs14-18. None of these deletions caused a major loss of synapses, suggesting that neuroligins are not essential for synapse formation, even though overexpression of neuroligins leads to an apparent increase in synapse numbers26 that has led many investigators to postulate that neuroligins function in ‘making’ synapses.
Neuroligin deletions in vivo, analyzed in multiple systems (Schaffer-collateral synapses of the CA1 region of the hippocampus, parallel-fiber and climbing-fiber synapses of the cerebellum, and calyx of Held synapses in the brain stem), produced a similar pattern of phenotypes as in neuroligin deletions in cultures with some exceptions19-25. The Nlgn3 deletion altered tonic endocannabinoid release at CCK-positive synapses in the CA1 region without altering other synapse properties24. In calyx of Held synapses, however, the same deletion altered the alignment of AMPAR nanoclusters, resulting in a decrease in synaptic strength25. The Nlgn1 deletion in the CA1 region, conversely, suppressed NMDAR EPSCs and blocked NMDAR-dependent LTP without altering AMPAR EPSCs20,21. Notably, the NMDAR EPSC phenotype could be rescued by Nlgn1 mutants unable to bind to neurexins but not with Nlgn1 mutants lacking a cytoplasmic tail, whereas the LTP phenotype exhibited the opposite Nlgn1 sequence requirements, suggesting that these two functions are mediated by distinct Nlgn1 interactions and mechanisms21. This conclusion is consistent with earlier work showing that neurexin binding-deficient Nlgn1 is still capable of inducing synapse growth, as is dimerization-deficient Nlgn126. The Nlgn4 deletion caused a significant suppression of inhibitory glycinergic synaptic transmission27. Finally and importantly, Nlgn3 and Nlgn4 missense mutations observed in severe cases of autism produce a gain-of-function phenotype as shown in vivo in mice and in culture with human neurons using knockin mutations17,18,23-25,28,29, although the gain-of-function mechanisms remain unclear.
Knowledge gaps. Again, despite a large number of papers (>1,100 papers in PubMed as of November 2023), neuroligins are poorly understood. Among the many key questions are i) what mechanisms render Nlgn1 and Nlgn2 specific for excitatory and inhibitory synapses, respectively; ii) how do various neuroligins perform synapse-specific functions, such as the role of Nlgn3 in tonic endocannabinoid secretion in CA124 or the role of Nlgn1 in NMDAR-dependent LTP20,21; iii) what other ligands mediate neuroligin functions since current neuroligin ligands (neurexins and MDGAs) do not account for all of their functions; iv) why do deletions of neuroligins have no effect on synapse numbers or size even though overexpression causes synapse growth; and v) what is the mechanism of the gain-of-function effects of autism-associated Nlgn3 and Nlgn4 mutations and do these mechanisms potentially provide insight into autism pathogenesis? Addressing these questions will be essential for progress in understanding the dynamic molecular logic of synaptic connections in neural circuits.
References
1. Ichtchenko, K., Hata, Y., Nguyen, T., Ullrich, B., Missler, M., Moomaw, C., Südhof, T.C. (1995) Neuroligin 1: A splice-site specific ligand for ß-neurexins. Cell 81, 435-443.
2. Ichtchenko, K., Nguyen, T., Südhof, T.C. (1996) Structures, alternative splicing, and neurexin binding of multiple neuroligins. J. Biol. Chem. 271, 2676-2682.
3. Bolliger, M.F., Pei, J., Maxeiner, S., Boucard, A.A., Grishin, N.V. Südhof, T.C. (2008) Unusually Rapid Evolution of Neuroligin-4 in Mice. Proc. Natl. Acad. Sci. U.S.A. 105, 6421-6426.
4. Maxeiner, S., Benseler, F., Krasteva-Christ, G., Brose, N., Südhof, T.C. (2020) Evolution of the Autism-Associated Neuroligin-4 Gene Reveals Broad Erosion of Pseudoautosomal Regions in Rodents. Molecular Biology and Evolution 37, 1243-1258.
5. Qin L, Guo S, Han Y, Wang X, Zhang B. (2020) Functional mosaic organization of neuroligins in neuronal circuits. Cell Mol Life Sci. 77, 3117-3127.
6. Südhof, T.C. (2017) Synaptic Neurexin Complexes: A Molecular Code for the Logic of Neural Circuits. Cell 171, 745-769.
7. Südhof, T.C. (2008) Neuroligins and Neurexins Link Synaptic Function to Cognitive Disease. Nature 455, 903-911.
8. Golf, S,R,, Trotter, J,H,, Nakahara, G., Südhof, T.C. (2023) Astrocytic Neuroligins Are Not Required for Synapse Formation or a Normal Astrocyte Cytoarchitecture. bioRxiv 10:2023.04.10.536254. doi: 10.1101/2023.04.10.536254.
9. Connor SA, Elegheert J, Xie Y, Craig AM (2019) Pumping the brakes: suppression of synapse development by MDGA-neuroligin interactions. Curr. Opin. Neurobiol. 57, 71-80.
10. Comoletti, D., Flynn, R.E., Boucard, A.A., Demeler, B., Schirf, V., Shi, J., Jennings, L.L., Newlin, H.R., Südhof, T.C., Taylor, P. (2006) Gene Selection, Alternative Splicing, and Post-Translational Processing Regulate Neuroligin Selectivity for b-Neurexins. Biochemistry 45, 12816-12827.
11. Boucard, A.A., Chubykin, A.A., Comoletti, D., Taylor, P., Südhof, T.C. (2005) A Splice-Code for Trans-Synaptic Cell Adhesion Mediated by Binding of Neuroligin 1 to a- and b-Neurexins. Neuron 48, 229-236.
12. Wang, C.Y., Trotter, J.H., Liakath-Ali, K., Lee, S.J., Liu, X., Südhof, T.C. (2021) Molecular Self-Avoidance in Synaptic Neurexin Complexes. Science Advances 7, eabk1924.
13. Yoshida, T., et al. (2021) Canonical versus non-canonical transsynaptic signaling of neuroligin 3 tunes development of sociality in mice. Nat. Commun. 12, 1848.
14. Varoqueaux, F., Aramuni, G., Rawson, R.L., Mohrmann, R., Missler, M., Gottmann, K., Zhang, W., Südhof, T.C., Brose, N. (2006) Neuroligins Determine Synapse Maturation and Function. Neuron 51, 741-754.Varoqueaux
15. Chubykin, A.A., Atasoy, D., Etherton, M.R., Brose, B., Kavalali, E.T., Gibson, J.R., Südhof, T.C. (2007) Activity-Dependent Validation of Excitatory vs. Inhibitory Synapses by Neuroligin-1 vs. Neuroligin-2. Neuron 54, 919-931.
16. Chanda, S., Hale, D.W., Zhang, B., Wernig, M., and Südhof, T.C. (2017) Unique vs. Redundant Functions of Neuroligin Genes in Shaping Excitatory and Inhibitory Synapse Properties. J. Neurosci. 37, 6816-6836.
17. Wang, L., Mirabella, V., Dai, R., Su, X., Xu, R., Jadali, A., Bernabucci, M., Singh, I., Chen, Y. Tian, J., Jiang, P., Kwan, K., Pak, C.H., Liu, C., Comoletti, D., Hart, R., Chen, C., Südhof, T.C., Pang, Z. (2022) Analyses of the Autism-associated Neuroligin-3 R451C Mutation in Human Neurons Reveals a Gain-of-Function Synaptic Mechanism. Mol. Psychiatry in press.
18. Marro, S., Chanda, S., Yang, N., Janas, J.A., Valperga, G., Trotter, J.H., Zhou, B., Merrill, S., Yousif, I., Shelby, H., Vogel, H., Kalani, M.Y.S., Südhof, T.C., Wernig, M. (2019) Neuroligin-4 regulates excitatory synaptic transmission in human neurons. Neuron 103, 617-626.
19. Zhang, B., Chen, L.Y., Liu, X., Maxeiner, S., Lee, S.J., Gokce, O., Südhof, T.C. (2015) Neuroligins Sculpt Cerebellar Purkinje-Cell Circuits by Differential Control of Distinct Classes of Synapses. Neuron 87, 781-796.
20. Jiang, M., Polepalli, J., Chen, L.Y., Zhang, B., Südhof, T.C., Malenka, R.C. (2016) Conditional ablation of neuroligin-1 in CA1 pyramidal neurons blocks LTP by a cell-autonomous NMDA receptor-independent mechanism. Mol. Psychiatry 22, 375-383.
21. Wu, X., Morishita, W.K., Riley, A.M., Hale, W.D., Südhof, T.C., Malenka, R.C. (2019) Neuroligin-1 Signaling Controls LTP and NMDA-Receptors by Distinct Molecular Pathway. Neuron 102, 621-635.
22. Zhang, B., Seigneur, E., Wei, P., Gokce, O., Morgan, J., Südhof, T.C. (2017) Developmental plasticity shapes synaptic phenotypes of autism-associated neuroligin-3 mutations in the calyx of Held. Mol. Psychiatry 22, 1483-1491.
23. Tabuchi, K., Blundell, J., Etherton, M.R., Hammer, R.E., Liu, X., Powell, C.M., Südhof, T.C. (2007) A Neuroligin-3 Mutation Implicated in Autism Increases Inhibitory Synaptic Transmission in Mice. Science 318, 71-76.
24. Földy, C., Malenka, R.C., Südhof, T.C. (2013) Autism-Associated Neuroligin-3 Mutations Commonly Disrupt Tonic Endocannabinoid Signaling. Neuron 78, 498-509.
25. Han, Y., Cao, R., Qin, L., Chen, L.Y., Tang, A.H., Südhof, T.C., Zhang, B. (2022) Neuroligin-3 confines AMPA-receptors into nanoclusters, thereby controlling synaptic strength at the calyx of Held synapses. Science Advances, 17, eabo4173.
26. Ko, J., Zhang, C., Araç, D., Boucard, A., Brunger, A.T., Südhof, T.C. (2009) Neuroligin-1 performs neurexin-dependent and neurexin-independent functions in synapse validation. EMBO J. 28, 3244-3255.
27. Zhang, B., Hale, D., Brose, N., Südhof, T.C. (2018) Autism-Associated Neuroligin-4 Mutation Selectively Impairs Glycinergic Synaptic Transmission in Mouse Brainstem Synapses. J. Exp. Medicine 215, 1543-1553.
28. Etherton, M.R., Földy C, Sharma, M., Tabuchi, K., Liu, X., Shamloo, M., Malenka, R.C., Südhof, T.C. (2011) Autism-linked neuroligin-3 R451C mutation differentially alters hippocampal and cortical synaptic function. Proc. Natl. Acad. Sci. U.S.A. 108, 13764-13769.
29. Rothwell, P.E., Fuccillo, M.V., Maxeiner, S., Hayton, S.J., Gokce, O., Lim, B.K., Fowler, S.C., Malenka, R.C., Südhof, T.C. (2014) Autism-Associated Neuroligin-3 Mutations Commonly Impair Striatal Circuits to Boost Repetitive Behaviors. Cell 158, 198-212.
Neurexin-LRRTM1/2 Complexes
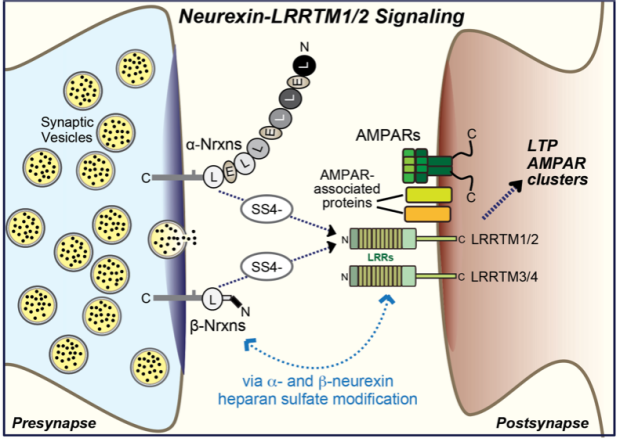
Properties. LRRTMs constitute a family of leucine-rich repeat proteins encoded by four genes (LRRTM1-4 in humans, Lrrtm1-4 in mice) that are primarily expressed in neurons1. LRRTMs are type 1 membrane proteins with a simple domain structure composed of an extracellular LRR-domain containing 10 leucine-rich repeats flanked by canonical N- and C-terminal cap sequences, a single transmembrane region, and a short cytoplasmic tail. Remarkably, all LRRTM genes except for the LRRTM4 gene include a single coding exon and are located in introns of different alpha-catenin genes, although no functional relation of LRRTMs to alpha-catenins is known1.
Interactions. LRRTM1 and LRRTM2, and possibly also LRRTM3, bind to SS4- but not to SS4+ neurexins2-4. A crystal structure of the LRRTM2-Nrxn1b complex revealed that the neurexin LNS6 domain is nestled into the C-terminal end of the LRR domain5. LRRTM4 binds to heparan-sulfate modified glypican-46, and LRRTM3 and LRRTM4 both bind to the heparan-sulfate moieties of neurexins, suggesting that LRRTM3 and LRRTM4 but not LRRTM1 and LRRTM2 are heparan-sulfate receptors7. Strikingly, LRRTM4 was also identified as part of a large multiprotein complex associated with AMPARs that may also contain other LRRTMs8. How all of these interactions fit together remains unclear.
Function. LRRTMs likely perform multiple synaptic functions. The best established is a redundant role of LRRTM1 and LRRTM2 in enabling NMDAR-dependent LTP and in regulating AMPARs9,10. These two functions may be connected and depend on binding of LRRTMs to AMPAR complexes, as suggested by their role in controlling the subsynaptic AMPAR position11.
Knowledge gaps. Again many of the fundamentals are still unclear.
References
1. Laurén J, Airaksinen MS, Saarma M, Timmusk T. (2003) A novel gene family encoding leucine-rich repeat transmembrane proteins differentially expressed in the nervous system. Genomics 81, 411-421.
2. Ko, J., Fuccillo, M., Malenka, R.C., and Südhof, T.C. (2009) LRRTM2 Functions as a Neurexin Ligand in Promoting Excitatory Synapse Formation. Neuron 64, 791-798.
3. de Wit J, Sylwestrak E, O'Sullivan ML, Otto S, Tiglio K, Savas JN, Yates JR 3rd, Comoletti D, Taylor P, Ghosh A. (2009) LRRTM2 interacts with Neurexin1 and regulates excitatory synapse formation. Neuron 64, 799-806.
4. Um JW, Choi TY, Kang H, Cho YS, Choii G, Uvarov P, Park D, Jeong D, Jeon S, Lee D, Kim H, Lee SH, Bae YC, Choi SY, Airaksinen MS, Ko J. (2016) LRRTM3 Regulates Excitatory Synapse Development through Alternative Splicing and Neurexin Binding. Cell Rep. 14, 808-822.
5. Yamagata A, Goto-Ito S, Sato Y, Shiroshima T, Maeda A, Watanabe M, Saitoh T, Maenaka K, Terada T, Yoshida T, Uemura T, Fukai S. (2018) Structural insights into modulation and selectivity of transsynaptic neurexin-LRRTM interaction. Nat. Commun. 9, 3964.
6. de Wit J, O'Sullivan ML, Savas JN, Condomitti G, Caccese MC, Vennekens KM, Yates JR 3rd, Ghosh A. (2013) Unbiased discovery of glypican as a receptor for LRRTM4 in regulating excitatory synapse development. Neuron 79, 696-711.
7. Roppongi RT, Dhume SH, Padmanabhan N, Silwal P, Zahra N, Karimi B, Bomkamp C, Patil CS, Champagne-Jorgensen K, Twilley RE, Zhang P, Jackson MF, Siddiqui TJ. (2020) LRRTMs Organize Synapses through Differential Engagement of Neurexin and PTPσ. Neuron 106, 108-125.
8. Schwenk J, Harmel N, Brechet A, Zolles G, Berkefeld H, Müller CS, Bildl W, Baehrens D, Hüber B, Kulik A, Klöcker N, Schulte U, Fakler B. (2012) High-resolution proteomics unravel architecture and molecular diversity of native AMPA receptor complexes. Neuron 74, 621-633.
9. Soler-Llavina, G.J., Fuccillo, M.V., Ko, J., Südhof, T.C., and Malenka, R.C. (2011) The neurexin ligands, neuroligins and LRRTMs, perform convergent and divergent synaptic functions in vivo. Proc. Natl. Acad. Sci. U.S.A. 108, 16502-16509.
10. Bhouri, M., Morishita, W., Temkin, P., Goswami, D., Kawabe, H., Brose, N., Südhof, T.C., Craig, A.M., Siddiqui, T.J., and Malenka, R.C. (2018) Deletion of LRRTM1 and LRRTM2 in adult mice impairs basal AMPA receptor transmission and LTP in hippocampal CA1 pyramidal neurons. Proc. Natl. Acad. Sci. U.S.A. 115, E5382-E5389.
11. Ramsey AM, Tang AH, LeGates TA, Gou XZ, Carbone BE, Thompson SM, Biederer T, Blanpied TA (2021) Subsynaptic positioning of AMPARs by LRRTM2 controls synaptic strength. Sci. Adv. 7, eabf3126.
Neurexin-Cerebellin Complexes and Their Receptors
Properties. Cerebellins are C1q-like domain proteins composed of an N-terminal cysteine-rich domain and a C-terminal C1q-domain. The C1q-domains of cerebellins form obligatory trimers that in turn are dimerized via their N-terminal cysteine-rich domain, resulting in a dimer of trimers (see Figure)1. Of the four mammalian cerebellin genes (Cbln1-4 in mice and CBLN1-4 in humans), Cbln1 is expressed at high levels in granule cells of the mature cerebellum (hence its name) and at lower levels in distinct cell types in a few selected brain regions2-4, while Cbln2 is expressed in cerebellar granule cells early during development when its levels in cerebellum supersede those of Cbln1, but in adult is primarily produced in a selected subset of neurons in distinct brain regions, such as the medial habenula, the hippocampal subiculum, and the brainstem2,3. Cbln3 is expressed only in cerebellar granule cells and unable to assemble into homomultimers but requires Cbln1 or Cbln2 for export to the cell surface5. Cbln4, finally, is not detectably present in the cerebellum but highly expressed in a few specific cell types, such as pyramidal cells in the entorhinal cortex, medial habenula neurons, and interneurons of the hippocampal CA1 region and the prefrontal cortex2,6. Interestingly, although the medial habenula expresses high concentrations of both Cbln2 and Cbln4, these are present in distinct neuronal populations projecting to the central and lateral part of the interpeduncular nucleus via the fasciculus retroflexus7.
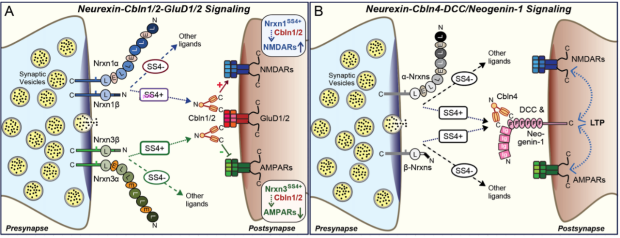
Interactions. Presynaptically, Cbln1, Cbln2, and Cbln4 bind to the LNS6 domain of a- and b-neurexins, but only when this domain contains an insert in SS4 (see Figure; note that Cbln3 has not been tested in any interactions)1. The structural mechanism of neurexin-binding by Cbln1 and Cbln2 is unknown but involves their cysteine-rich N-terminal ‘stalk’. Postsynaptically, Cbln1 and Cbln2 bind to the N-terminal domains of GluD1 and GluD2, homologs of AMPARs and NMDARs that probably do not function as ligand-gated ionotropic receptors8. Cbln4, in turn, bind to postsynaptic DCC and neogenin-1 (which are highly homologous Ig- and FNIII-domain proteins, but does not bind to GluD’s with high affinity6,9-11. All cerebellins bind to the postsynaptic receptors via their trimeric C1q-like domains which thus exhibit interaction specificity for distinct targets despite being highly homologous.
Functions. The loss of parallel-fiber synapses in Cbln1 KO mice (<50%) and the observation of ‘naked’ Purkinje cell spines (<5%) in Cbln1 KO mice prompted the hypothesis that cerebellins mediate synapse formation12. However, parallel-fiber synapses are initially formed in Cbln1 KO mice but then seem to degenerate. Moreover, the Cbln1 KO produces a loss of LTD in all parallel-fiber synapses but only the loss of a subset of parallel-fiber synapses12. Indeed, constitutive double deletions of Cbln1 and Cbln2 and triple deletions of Cbln1, Cbln2, and Cbln4 causes seizures and motor behavior impairments but do not decrease synapse numbers in the forebrain of young adult mice, although these deletion do decrease the abundance of some synapse types in older mice7,13. These results suggest that at least in the forebrain, cerebellins are not essential for synapse formation but are necessary for the long-term stabilization of synapses.
Deletions of cerebellins do, however, produce robust functional synaptic phenotypes that manifest quickly after a conditional deletion is induced. These phenotypes differ depending on the specific synapses that are being studied, as first shown in the loss of LTD at parallel-fiber synapses induced by the Cbln1 KO12. From studies of our lab, two key results stand out.
First, deletion of Cbln2 in the hippocampal subiculum causes an upregulation of AMPAR-EPSCs and a down-regulation of NMDAR-EPSCs without changing synapse numbers14. These phenotypes were replicated by the GluD2 deletion. Strikingly, ablating expression of presynaptic Nrxn3-SS4+ variants downregulated AMPARs without affecting NMDARs, whereas ablating expression of presynaptic Nrxn1-SS4+ variants upregulated NMDARs without affecting AMPARs14. Thus, the combination of the Nrxn1-SS4+ and Nrxn3-SS4+ ablation is a mirror image of the Cbln2 and GluD2 deletions (see Figure). Deletion of Cbln2 in the mPFC, conversely, only affected NMDARs without changing AMPARs at excitatory synapses, suggesting that different types of synapses respond differentially to the Cbln2-GluD2 signaling pathway15.
Second, deletion of Cbln4 in the entorhinal cortex produced a loss of LTP in the perforant-path synapses formed on dentate gyrus granule cells without changing either synapse numbers or basal synaptic strength6. A postsynaptic deletion of the Cbln4 receptor neogenin1 in the dentate gyrus induced the same phenotype6, suggesting that the neurexin-Cbln4-neogenin1 signaling complex organizes perforant-path synapses to render them LTP-competent.
Knowledge gaps. Neurexin-cerebellin complexes are surrounded by fundamental questions, including the following: First, does the secondary loss of some synapses upon the deletion of cerebelllins signify a stabilization function of cerebellins for subsets of synapses or are they the non-specific downstream effects of a change in synapse properties? Second, what intracellular signaling mechanisms mediate the synapse-organizing functions of cerebellins? Third, at a more basic level, what is the structure of neurexin-Cbln1/2-GluD or neurexin-Cbln4-neogenin/DCC complexes, and do cerebellins heteromultimerize? Fourth, how does a presynaptic Nrxn1 vs. Nrxn3 signal instruct postsynaptic GluD’s to regulate NMDAR- vs. AMPAR-receptors via the same Cbln1/2 signaling intermediate? Fifth, given that at least Cbln2 and Cbln4 expression may be activity-dependent, do cerebellins mediate synapse restructuring as a function of activity? Sixth, in view of the striking neuron type-specific expression patterns of various cerebellins, is there a circuit-specific function of various cerebellins as suggested by the role of Cbln2 in controlling compulsive behaviors?16 Finally, do cerebellins have a role in neuropsychiatric disorders? Again, much work remains to be done.
References
1. Yuzaki, M. (2018) Two Classes of Secreted Synaptic Organizers in the Central Nervous System. Annu. Rev. Physiol. 80, 243-262.
2. Miura E, Iijima T, Yuzaki M, Watanabe M. Distinct expression of Cbln family mRNAs in developing and adult mouse brains.Eur J Neurosci. 2006 Aug;24(3):750-60.
3. Seigneur, E., Südhof, T. C. (2017) Cerebellins are differentially expressed in selective subsets of neurons throughout the brain. J. Comp. Neurol. 525, 3286-3311.
4. Wang, C.Y., Trotter, J.H., Liakath-Ali, K., Lee, S.J., Liu, X., Südhof, T.C. (2021) Molecular Self-Avoidance in Synaptic Neurexin Complexes. Science Advances 7, eabk1924.
5. Pang, Z., Zuo, J., Morgan, J.I. (2000) Cbln3, a novel member of the precerebellin family that binds specifically to Cbln1.J. Neurosci. 20, 6333-6339.
6. Liakath-Ali, K., Polepalli, J.S., Lee, S.J., Cloutier, J.F., Südhof, T.C. (2022) Transsynaptic Cerebellin 4-Neogenin 1 Signaling Mediates LTP in the Mouse Dentate Gyrus. Proc. Natl. Acad. Sci. U.S.A. 119, e2123421119.
7. Seigneur, E., Pollepali, J., Südhof, T.C. (2018) Cbln2 and Cbln4 are expressed in distinct medial habenula-interpeduncular projections and contribute to different behavioral outputs. Proc. Natl. Acad. Sci. U.S.A. 115, E10235-E10244.
8. Yuzaki, M., Aricescu, A.R. (2017) A GluD Coming-Of-Age Story. Trends Neurosci. 40, 138-150.
9. Wei, P., Pattarini, R., Rong, Y., Guo, H., Bansal, P.K., Kusnoor, S.V., Deutch, A.Y., Parris, J., Morgan, J.I. (2012) The Cbln family of proteins interact with multiple signaling pathways.J Neurochem. 121, 717-729.
10. Zhong C, Shen J, Zhang H, Li G, Shen S, Wang F, Hu K, Cao L, He Y, Ding J. (2017) Cbln1 and Cbln4 Are Structurally Similar but Differ in GluD2 Binding Interactions. Cell Rep. 20, 2328-2340.
11. Cheng, S., Seven, A.B., Wang, J., Skiniotis, G., Özkan, E. (2016) Conformational Plasticity in the Transsynaptic Neurexin-Cerebellin-Glutamate Receptor Adhesion Complex. Structure 24, 2163-2173.
12. Hirai, H., Pang, Z., Bao, D., Miyazaki, T., Li, L., Miura, E., Parris, J., Rong, Y., Watanabe, M., Yuzaki, M., Morgan, J.I. (2005) Cbln1 is essential for synaptic integrity and plasticity in the cerebellum. Nat. Neurosci. 8, 1534-1541.
13. Seigneur, E., Südhof, T. C. (2018) Genetic ablation of all cerebellins reveals synapse organizer functions in multiple regions throughout the brain. J. Neurosci. 38, 4774-4790.
14. Dai, J., Patzke, C., Liakath-Ali, K., Seigneur, E., Südhof, T.C. (2021) GluD1, A signal transduction machine disguised as an ionotropic receptor. Nature 595, 261-265.
15. Dai, J., Liakath-Ali, K., Golf, S., Südhof, T.C. (2022) Distinct Neurexin-Cerebellin Complexes Control AMPA- and NMDA-Receptor Responses in a Circuit-Dependent Manner. E-Life 11, e78649.
16. Seigneur, E., Polepalli, J., Wang, J., Dai, J., Südhof, T.C. (2021) Cerebellin-2 Regulates a Serotonergic Dorsal Raphe Circuit that Controls Compulsive Behaviors. Mol. Psychiatry. 26, 7509-7521.
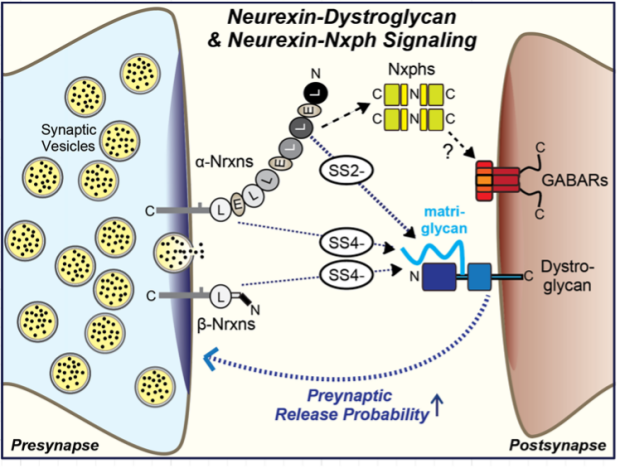
Neurexin-Dystroglycan Complexes
Properties. Dystroglycan is a broadly expressed cell-adhesion molecule that contains a unique glycosaminoglycan-like polysaccharide modification called ‘matriglycan’ that is largely composed of a glucuronic acid-β1,3-xylose disaccharide repeat1. Dystroglycan performs an array of functions in and outside of brain via the binding of the Matrigel moiety to the LNS-domains (also called LG-domains) of multiple proteins, including the LNS2 and LNS6 domains of neurexins2. Intracellularly, dystroglycan binds to dystrophins and related molecules to link cell adhesion to the cytoskeleton. Impairments of the dystroglycan protein complex by mutations in dystrophin or in the enzymes generating the unique matriglycan modification of dystroglycan cause muscular dystrophy and cognitive impairments.
Interactions. The atomic structure of the matriglycan moiety bound to laminin LG/LNS-domains revealed that negative charges from the matriglycan’s glucuronic acid complete the coordination spheres of a Ca2+ ion of the LG/LNS-domain, thereby achieving a high binding affinity3. In the case of neurexins, only LNS2 and LNS6 bind to dystroglycan, with LNS2 exhibiting tighter binding than LNS6. The mechanism of dystroglycan binding to neurexin LNS-domains is likely the same as for LNS-domains of laminin and many other proteins. Neurexin LNS2 and LNS6 are alternatively spliced at similar positions at SS2 and SS4; in both cases, only the minus-insert variants bind to neurexins2,4.
Function. Dystroglycan performs an essential function at a subset of inhibitory synapses that was initially thought to be independent of neurexin binding because of indirect data5. However, we recently showed that the deletions of presynaptic Nrxn3 and of postsynaptic dystroglycan cause identical phenotypes in olfactory bulb granule cellmitral cell synapses and in prefrontal cortex inhibitory synapses6. Moreover, we found that the Nrxn3 deletion phenotypes can be selectively rescued with an Nrxn3 construct containing only the LNS2-SS2- domain without any other LNS or EGF-like domains but cannot be rescued with the equivalent LSN2-SS2+ domain construct6. Strikingly, the phenotype of the dystroglycan and Nrxn3 deletions consists of a decrease in release probability, suggesting that presynaptic Nrxn3 controls the presynaptic release probability via a mechanism that requires the trans-synaptic interaction of presynaptic Nrxn3 with postsynaptic dystroglycan (see Figure).
Knowledge gaps. The mechanism by which the neurexin-dystroglycan interaction regulates the presynaptic release probability is unclear. It is also unknown whether dystroglycan physiologically interacts with Nrxn1 and Nrxn2 in addition to Nrxn3. Moreover, it has not yet been clarified if dystroglycan functions via an interaction with neurexins in other synapses, for example at inhibitory cerebellar synapses in which dystroglycan plays a central role7.
References
1. Barresi, R., Campbell, K.P. (2006) Dystroglycan: from biosynthesis to pathogenesis of human disease. J. Cell Sci. 119 (Pt 2), 199-207.
2. Sugita, S., Saito, F., Tang, J., Satz, J., Campbell, K., Südhof, T.C. (2001) A stoichiometric complex of neurexins and dystroglycan in brain. J. Cell Biol. 154, 435-445.
3. Briggs, D.C., Yoshida-Moriguchi, T., Zheng, T., Venzke, D., Anderson, M.E., Strazzulli, A., Moracci, M., Yu, L., Hohenester, E., Campbell, K.P. (2016) Structural basis of laminin binding to the LARGE glycans on dystroglycan. Nat. Chem. Biol. 12, 810-814.
4. Reissner, C., Stahn, J., Breuer, D., Klose, M., Pohlentz, G., Mormann, M., Missler, M. (2014) Dystroglycan binding to a-neurexin competes with neurexophilin-1 and neuroligin in the brain. J. Biol. Chem 289, 27585 – 27603.
5. Früh, S., Romanos, J., Panzanelli, P., Bürgisser, D., Tyagarajan, S.K., Campbell, K.P., Santello, M., Fritschy, J.M. (2016) Neuronal Dystroglycan Is Necessary for Formation and Maintenance of Functional CCK-Positive Basket Cell Terminals on Pyramidal Cells. J. Neurosci. 36, 10296-10313.
6. Trotter, J.H., Wang, C.Y., Zhou, P., Nakahara, G., Südhof, T.C. (2023) A combinatorial code of neurexin-3 alternative splicing controls inhibitory synapses via a trans-synaptic dystroglycan signaling loop. Nature Comm. 14, 1771.
7. Briatore, F., Pregno, G., Di Angelantonio, S., Frola, E., De Stefano, M.E., Vaillend, C., Sassoè-Pognetto, M., Patrizi, A. (2020) Dystroglycan Mediates Clustering of Essential GABAergic Components in Cerebellar Purkinje Cells. Front. Mol. Neurosci. 13, 164.
Neurexin-Neurexophilin Complexes
Properties. Neurexophilins are a family of secreted neurexin-ligands encoded by four genes (NXPH1-4 in humans, Nxph1-4 in mice) that exhibit a ‘cytokine-like’ pattern of disulfide-bonded cysteine residues but are not homologous to any other protein in databases1,2. Neurexophilins are primarily, if not exclusively, expressed in neurons in a differential pattern. Nxph1 is present at modest levels in inhibitory interneurons in forebrain but also in discrete neuronal cell types in the brainstem, Nxph2 is barely detectable in brain but highly expressed in a subset of cholinergic interneurons of the gut, Nxph3 is abundant in a small subset of excitatory cortical neurons, and Nxph4 is primarily expressed in subsets of forebrain interneurons. All neurexophilins contain a signal peptide, a putative propeptide of approximately 57-82 amino acids that ends in a dibasic sequence motif, and two larger blocks of highly conserved sequences (72-75 and 84-86 amino acids) that are separated by a variable non-conserved sequence of 9-57 residues.
Interactions. Neurexophilins have only one validated interaction, their binding to the LNS2 domain of a-neurexins2,3. All a-neurexins and neurexophilins bind to each other. Their interaction is modulated by alternative splicing at SS2 in the LNS2 domain, such that Nxph1 binds to Nrxn1-SS2- variants with an approximately 6-fold lower affinity than to Nrxn1-SS2+ variants (apparent Kd’s ~300 nM and 50 nM, respectively)4. Since dystroglycan also binds to neurexin LNS2 domains but only to their SS2- splice variants5, neurexophilins and dystroglycan compete with each other for binding to neurexins, with neurexophilins exhibiting the higher apparent affinity in vitro6.
The structure of the Nrxn1 LNS2-SS2+ and LNS2-SS2- complexes with Nphn1 lacking the putative propeptide revealed that the two conserved blocks of Nphn1 sequences form a single continuous jelly-roll b- sandwich that seamlessly extends into the jelly-roll b-sandwich formed by the Nrxn1 LNS2 domain, with a unique, extensive interface connecting the two jelly-roll b-sandwiches4. The SS2A insert of LNS2 augments this interface, increasing the binding affinity of the Nrxn1 LNS2 domain for Nphn1 (see Figure taken from ref. 4).
Nxph4 is co-immunoprecipitated from brain not only with neurexins but also with GABA-receptors, suggesting an interaction7 although it is unclear whether this is a direct or indirect interaction since neurexins have also been shown to bind to GABA-receptors8.
Function. Unknown. A Nxph4 KO severely impairs (>80%) the Golgi cellgranule cell synaptic connectivity in the cerebellum but produces only an ~10% loss of the number of these synapses, suggesting that is important for the function and not the formation of cerebellar Golgi cellgranule cell synapses7.
Knowledge gaps. Only the structures, expression patterns, and neurexin-binding of neurexophilins are known. Key questions now include the relative affinities and functions of different neurexophilins and their relation to dystroglycan binding. Maybe the most important question, however, is what neurexophilins do and how they do it. For example, it is unknown if neurexophilins are simply neurexin agonists that stimulate the function of neurexins engaged simultaneously in other interactions (see Figure), or whether they serve as adaptors that connect neurexins to a unidentified postsynaptic receptor. Given the strikingly differentiated expressing patterns of neurexophilins, exploring their functions in particular in defined classes of interneurons that express high levels of a given isoform but also use dystroglycan as a synaptic adhesion molecule would be rewarding.
References
1. Petrenko, A.G., Ullrich, B., Missler, M., Krasnoperov, V., Rosahl, T.W., Südhof, T.C. (1996) Structure and evolution of neurexophilin. J. Neurosci. 16, 4360-4369.
2. Missler, M., and Südhof, T. C. (1998) Neurexophilins form a conserved family of neuropeptide-like glycoproteins. J. Neurosci. 18, 3630-3638.
3. Missler, M., Hammer, R.E., Südhof, T.C. (1998) Neurexophilin binding to α-neurexins: A single LNS-domain functions as independently folding ligand-binding unit. J. Biol. Chem. 273, 34716-34723.
4. Wilson, S., White, K.I., Zhou, Q., Pfuetzner, R.A., Choi, U.B., Südhof, T.C., Brunger, A.T. (2019) Structures of Neurexophilin-Neurexin Complexes Reveal a Regulatory Mechanism of Alternative Splicing. EMBO J. 38: e101603.
5. Sugita, S., Saito, F., Tang, J., Satz, J., Campbell, K., Südhof, T.C. (2001) A stoichiometric complex of neurexins and dystroglycan in brain. J. Cell Biol. 154, 435-445.
6. Reissner, C., Stahn, J., Breuer, D., Klose, M., Pohlentz, G., Mormann, M., Missler, M. (2014) Dystroglycan binding to a-neurexin competes with neurexophilin-1 and neuroligin in the brain. J. Biol. Chem 289, 27585 – 27603.
7. Meng, X., McGraw, C.M., Wang, W., Jing, J., Yeh, S.Y., Wang, L., Lopez, J., Brown, A.M., Lin, T., Chen, W., Xue, M., Sillitoe, R.V., Jiang, X., Zoghbi, H.Y. (2019) Neurexophilin4 is a selectively expressed α-neurexin ligand that modulates specific cerebellar synapses and motor functions. Elife 8, e46773.
8. Zhang, C., Atasoy, D., Araç, D., Yang, X., Fuccillo, M.V., Robison, A.J., Ko, J., Brunger, A.T., and Südhof, T.C. (2010) Neurexins Physically and Functionally Interact with GABAA-receptors. Neuron 66, 403-416.
Neurexin Cis-Complexes with LAR-Type PTPRs and the Heparan Sulfate Enigma
Neurexins, including Nrxn1g that contains only the juxtamembranous ‘cys-loop’ domain (see Figure above), are among several synaptic molecules that are modified by heparan sulfate, which include glypicans and syndecans1-3. Neurexins bind to multiple other synaptic adhesion molecules via their heparan sulfate modification, most prominently LAR-type receptor phosphotyrosine phosphatases (LAR-PTPRs)4,5. Corresponding to the large number of synaptic heparan sulfate-modified proteins, many synaptic haparan sulfate-binding proteins were described, including but not limited to LAR-PTPRs, LRRTM3 and LRRTM4, Unc5, and neuroligin-1. The neurexin-LAR-PTPR interaction occurs in cis in contrast to the neurexin-LRRTM4 interaction in trans that is also mediated by the neurexin heparan sulfate modification (see discussion above).
The common properties of presynaptic neurexins and LAR-PTPRs as hub molecules interacting with a large number of trans-synaptic ligands raised the possibility that they may be functionally redundant. Indeed, when we analyzed the effect of conditional sextuple deletions of all neurexins (except for Nrxn1g) and of all LAR-PTPRs at synapses formed by Purkinje cell outputs onto deep cerebellar nuclei, we observed a nearly complete ablation of synaptic connections in the sextuple deletion that was not present in the triple deletions of either neurexins or LAR-PTPRs6. Thus, at least at a subset of synapses these two presynaptic adhesion molecules are functionally redundant.
Knowledge gaps. The functional redundancy between neurexins and LAR-PTPRs does not tell us whether their interaction is functionally important. Mutation of the Nrxn1 heparan sulfate modification sequence causes a mild synaptic phenotype consistent with a physiological role of this modification in synapses1 but this role might be mediated by other heparan sulfate-dependent interactions of Nrxn1. An even broader question is which of the many heparan sulfate-dependent synaptic interactions are specific, functionally relevant and occur in a physiological setting. Given the complexity of heparan sulfate modifications and of their promoted interactions, only direct measurements of affinities will address this important question.
References
1. Zhang P, Lu H, Peixoto RT, Pines MK, Ge Y, Oku S, Siddiqui TJ, Xie Y, Wu W, Archer-Hartmann S, Yoshida K, Tanaka KF, Aricescu AR, Azadi P, Gordon MD, Sabatini BL, Wong ROL, Craig AM. (2018) Heparan Sulfate Organizes Neuronal Synapses through Neurexin Partnerships. Cell 174, 1450-1464
2. Sterky, F.H., Trotter, J.H., Lee, S., Recktenwald, C.V., Du, X., Zhou, B., Zhou, P., Schwenk, J., Fakler, B., Südhof, T.C. (2017) The Carbonic Anhydrase-Related Protein CA10 Is An Evolutionarily Conserved Pan-Neurexin Ligand. Proc. Natl. Acad. Sci. U.S.A. 114, E1253-E1262.
3. Kamimura, K., Maeda, N. (2021) Glypicans and Heparan Sulfate in Synaptic Development, Neural Plasticity, and Neurological Disorders. Front. Neural Circuits 15, 595596.
4. Han KA, Kim YJ, Yoon TH, Kim H, Bae S, Um JW, Choi SY, Ko J (2020) LAR-RPTPs Directly Interact with Neurexins to Coordinate Bidirectional Assembly of Molecular Machineries. J. Neurosci. 40, 8438-8462.
5. Roppongi RT, Dhume SH, Padmanabhan N, Silwal P, Zahra N, Karimi B, Bomkamp C, Patil CS, Champagne-Jorgensen K, Twilley RE, Zhang P, Jackson MF, Siddiqui TJ (2020) LRRTMs Organize Synapses through Differential Engagement of Neurexin and PTPs. Neuron 106, 108-125.
6. Sclip A., Südhof, T.C. (2023) Combinatorial Expression of Neurexins and LAR-type Phosphotyrosine Phosphatase Receptors Instructs Assembly of a Cerebellar Circuit. Nature Comm. 14, 4976.
Neurexin-CA10/11 and Neurexin-FAM19 complexes blocking heparan sulfate modifications of neurexins
We identified two families of secreted neurexin-binding proteins, CA10/111 and FAM19A-D (a.k.a. ‘TAFA1-4’), that bind close to the heparan sulfate modification site in the ‘stalk’ region of neurexins separating their 6th LNS-domain from the transmembrane region2. CA10 and CA11 are largely composed of a carbonic anhydrase-like domain whereas FAM19A-D are cysteine-rich proteins resembling cytokines. Interestingly, FAM19’s bind to neurexins in the secretory pathway with formation of a disulfide bond and are thus genuine neurexin subunits2. The functions of these two protein families remain unknown. Interestingly, both appear to interfere with the heparan sulfate modification of neurexins, suggesting that they may act to regulate this modification2,3.
References
1. Sterky, F.H., Trotter, J.H., Lee, S., Recktenwald, C.V., Du, X., Zhou, B., Zhou, P., Schwenk, J., Fakler, B., and Südhof, T.C. (2017) The Carbonic Anhydrase-Related Protein CA10 Is An Evolutionarily Conserved Pan-Neurexin Ligand. Proc. Natl. Acad. Sci. U.S.A. 114, E1253-E1262.
2. Khajal, A.J., Sterky, F.H., Sclip, A., Schwenk, J., Brunger, A.T., Fakler, B., and Südhof, T.C. (2020) Deorphanizing FAM19A Proteins as Pan-Neurexin Ligands with an Unusual Biosynthetic Binding Mechanism. J. Cell Biol. 219, e202004164
3. Montoliu-Gaya L, Tietze D, Kaminski D, Mirgorodskaya E, Tietze AA, Sterky FH. (2021) CA10 regulates neurexin heparan sulfate addition via a direct binding in the secretory pathway. EMBO Rep. 22, e51349.
Neurexins and Neurexin Ligands in Neuropsychiatric Disorders
Neurexins and many of their extra- and intracellular ligands have been implicated in neuropsychiatric diseases by human genetics1. Especially deletions within the very large NRXN1 gene (nearly 2 MB in size) produce a strong genetic predisposition for schizophrenia, autism, Tourette syndrome, and mental disability1,2. Analysis of patient-derived human neurons with heterozygous NRXN1 deletions or of human neurons with engineered heterozygous NRXN1 deletions revealed a robust decrease in excitatory synaptic transmission, suggesting that heterozygous NRXN1 deletions are pathophysiologically significant3,4. No such phenotype was observed in identically prepared mouse neurons with heterozygous Nrxn1 deletions, suggesting that human neurons are particularly sensitive to a decrease in Nrxn1 levels4. In addition, the heterozygous human NRXN1 deletion produced an increase in the levels of CASK, a cytoplasmic neurexin-binding scaffold protein5. A major challenge now is to explore whey heterozygous NRXN1 deletions, but also mutations in neuroligins and other neurexin-associated synaptic proteins, predispose to neuropsychiatric disorders is they cause a general impairment in synapses, and why even the same mutations can lead to distinct clinical presentations.
References
1. Südhof, T.C. (2008) Neuroligins and Neurexins Link Synaptic Function to Cognitive Disease. Nature 455, 903-911.
2. Castronovo, P., Baccarin, M., Ricciardello, A., Picinelli, C., Tomaiuolo, P., Cucinotta, F., Frittoli, M., Lintas, C., Sacco, R., Persico, A.M. (2020) Phenotypic spectrum of NRXN1 mono- and bi-allelic deficiency: A systematic review. Clin. Genet. 97, 125-137.
3. Pak, C.H., Danko, T., Zhang, Y., Aoto, J., Anderson, G., Maxeiner, S., Yi, F., Wernig, M., Südhof, T.C. (2015) Human Neuropsychiatric Disease Modeling Using Conditional Deletion Reveals Synaptic Transmission Defects Caused by Heterozygous Mutations in NRXN1. Cell Stem Cell 17, 316-328.
4. Pak, C., Danko, T., Mirabella, V.R., Wang, J., Liu, Y., Vangipuram, M., Grieder, S., Zhang, X., Ward, T., Huang, Y.W.A., Jin, K., Dexheimer, P., Bardes, E., Mittelpunkt, A., Ma, J., McMachlan, M., Moore, J.C., Qu, P., Purmann, C., Dage, J.L., Swanson, B.J., Urban A.E., Aronow, B.J., Pang, Z.P., Levinson, D.F., Wernig, M., Südhof, T.C. (2021) Cross-Platform Validation of Neurotransmitter Release Impairments in Schizophrenia Patient-Derived NRXN1-Mutant Neurons. Proc. Natl. Acad. Sci. U.S.A. 118, e2025598118.
5. Hata, Y., Butz, S., and Südhof, T.C. (1996) CASK: A novel dlg/PSD95 homologue with an N-terminal CaM kinase domain identified by interaction with neurexins. J. Neurosci. 16, 2488-2494.